阅读:0
听报道
More, more, more.

撰文 | Krestrel
蜜蜂(honey bee)是广为人知的社会性昆虫,常常被人们用来描述辛勤的劳动者。一个典型的蜂群由一只蜂后(也称为蜂王)、少量的雄蜂和大量的工蜂组成。群居的蜜蜂在职责上有着严格的划分,蜂后和雄蜂直接参与有性生殖,负责传宗接代,而工蜂负责育幼、筑巢、觅食和防务。(此处响起《打工人之歌》)
为了维持蜂群在组成上的平衡,蜜蜂的繁殖受到严格的调控。其中,工蜂和蜂后是从受精卵孵化而来,均为雌性;而雄蜂是从未受精的卵孵化而来——也就是说,雄蜂没有爸爸。
通常来说,有性生殖产生的后代同时继承了父亲和母亲各贡献的一套遗传物质。无论是工蜂、蜂后还是人类,都具有两套染色体组,也就是“二倍体”(diploid)。不同的是,工蜂和蜂后有16对染色体(总共32条),而人类则具有23对染色体(46条)。而未受精卵孵化而来的雄蜂则只具有正常染色体的一半(总共16条),也就是“单倍体”(haploid,谁让它没有爸爸呢)。
虽然工蜂和蜂后具有完全相同的染色体,但却只有蜂后才有生殖功能。科学家们发现,工蜂的生殖功能被抑制了,原因主要出在食物上。原来,蜂后一直都是以蜂王浆(royal jelly)为食物,而工蜂只在出生后吃了三天蜂王浆,以后只能靠花粉和花蜜为食。蜂王浆中具有调节基因表达的物质,能让一直吃蜂王浆的蜂后发展出成熟的生殖系统。另外,蜂后还会分泌外激素(pheromone),进一步抑制工蜂的生殖能力。所以工蜂其实是生殖器官发育不完全的雌性蜜蜂。
不过,在蜂王长时间缺失的条件下,工蜂的卵巢可以激活。但它们仍然不具有交配的能力,只能产下未受精的卵,发育成雄蜂。这种单性生殖的现象也叫孤雌生殖。工蜂单性繁殖的后代都是雄性,因此称为产雄孤雌生殖(arrhenotokous parthenogenesis,arrhenotoky)。这些雄蜂可以与其它蜂群的蜂王交配,将携带的基因组传递下去。
在蜂群分群之前,工蜂会在蜂巢内建起4-10个“王台”(queen cell),里面的幼虫用蜂王浆喂养,差不多同时完成发育。一般来说,一个种群只保留一只蜂王,这些王台中发育出来的蜂后,最强壮的会成为新群主,其他蜂后都会它被杀死。新的蜂后产生后,就到了分群(reproductive swarming)的时候。老蜂后会带着部分工蜂离开,建立新群。
与此相对应的是,因为一出来就马上面临一场生死较量,越早爬出王台对候选的蜂王越有利。需要尽快成熟的蜂王发育是最快的(15-16天),工蜂其次(21天),雄蜂最慢(24天)。
和人类不同,蜜蜂虽有雌雄,但没有性染色体,其性别决定机制比较特殊,称为单倍二倍性(haplodiploidy),主要存在于膜翅目昆虫和一些其它节肢动物中。但因为昆虫的多样性极大,这种性别决定机制的昆虫种类占到所有动物的20%,所以也不算那么特殊啦。
那么,蜜蜂的性别到底是由什么来决定的呢?
从分子水平上看,蜜蜂基因组中有一个称为csd的位点,是控制蜜蜂性别的主要基因位点,全称是互补性别决定子(complimentary sex determiner)。在蜜蜂种群中,这个位点的基因大概有19个不同的版本,也就是19个等位基因。雄蜂是单倍体,自然只有一个csd基因。蜂王和工蜂是二倍体,就有两个csd基因,而且这两个csd基因的版本总是不同(称为杂合子),因为版本相同(称为纯合子)的蜜蜂在发育早期就会死亡。
因此,csd位点是决定蜜蜂性别的最上游信号:csd杂合,决定了是雌性;csd纯合,蜜蜂无法活下来;只有单个csd基因,决定了是雄性。
也有一些特殊的情况,例如,在高度近交的实验种群中,csd位点处于纯合状态的二倍体会发育成雄蜂,但这样的雄蜂要么自身不育,要么交配产生的三倍体后代不育。
这样的性别决定机制与蜜蜂的“真社会性”(eusociality)是相适应的。所谓真社会性,是指种群要同时拥有三个特征:繁殖分工、世代重叠(群体中成熟个体可以分为两个以上的世代,比如爷爷辈和爸爸辈同时在世)和合作育幼。
工蜂的利他行为可以用亲缘选择学说来解释。亲缘系数(relatedness,简写为r)是用来衡量个体间亲缘关系的一个量,定义为两个个体共有某个基因且两个拷贝都来自同一祖先(例如母子的共有基因来源于母亲的父亲,即孩子的外公;姊妹的共有基因来源于她们的母亲)的概率。拿工蜂来说,产卵的工蜂与其产下的单倍体雄蜂的亲缘系数r=0.5,因为雄蜂继承了其母亲一半的遗传物质,拥有其母亲某个基因的概率为1/2。
而工蜂与蜂后产下的雄蜂的亲缘系数只有r=0.25,因为两者都是蜂后的后代:雄蜂拥有蜂王的某个基因的概率是1/2,工蜂作为雄蜂的姐妹,也拥有这个基因的概率也是1/2(还可能是来自另一只雄蜂),因此两者共有的这个基因均来自蜂王的概率就是1/4。
但是,蜂群里不会只有一只工蜂产卵,某只工蜂与所有工蜂产的雄蜂的亲缘系数r≈0.125,这样的话,与只照料蜂王产的雄蜂相比,照料到其它工蜂产的雄蜂反而亏了——为什么要照料亲缘系数更低的雄蜂呢?当然r越高,越是“自己人”。因此,在普通的蜂群中,工蜂自身不产卵,甚至还会除掉其它工蜂产的卵。
听起来很损是不是?其实,工蜂的这种行为,是为了增加自身的广义适合度。所谓适合度,是产生存活后代的能力,而广义适合度的要求更低:不必自己产生后代,别人的后代包含和自己相同的基因也行。(我的心不禁一颤:多可爱的小生灵啊!)
然而,十多年前,南非的养蜂人发现自己养的蜂群被一些能自行产卵的工蜂“颠覆”了。更富戏剧性的是,这些奇怪的工蜂产生的后代和自己的基因组是一样的,也就相当于克隆了自己!这些蜜蜂叫做海角蜜蜂(cape honeybee),是西方蜜蜂(学名Apis mellifera)的一个亚种。它的拉丁文学名是Apis mellifera capensis,区别于一般养蜂业采用的亚种Apis mellifera scutellata。

图1 “黑化了,但没有完全黑化”。腹部黑色比重更大是分辨出海角蜜蜂的一个方便特征。
这两个亚种的基因组几乎完全同源。但是,本文开头所讲到的典型工蜂只可能产下单倍体的雄蜂,而海角蜜蜂却能独自产下二倍体卵,发育成工蜂,或者,倘若占到了王台,发育成蜂王——你没听错,由工蜂产下的蜂王!这种现象称为产雌孤雌生殖(thelytokous parthenogenesis,thelytoky)。
看到这里,你可能会想起《生化危机·灭绝》里面的场景:在爱丽丝和克隆的爱丽丝面前,更多克隆爱丽丝悬浮在营养液里,被乘放在一望无际的一堆格状排列的卵形容器中(图2)。你或许还记得在《生化危机·终章》中,衰老的爱丽西亚将自己的回忆留给了自己的克隆体——主角爱丽丝赋予她完整的“人格”,希望她代替自己生存下去。事实上,澳大利亚悉尼大学昆虫学家Benjamin Oldroyd也用“转世”(reincarnation,字面意义是“重新支配肉体”)来指这个无法交配的工蜂产下自己的蜂王克隆的过程。

图2 电影《生化危机·灭绝》中相关场景。
的确,过去30年间,数百万海角蜜蜂克隆大军曾让南非养蜂业闻风丧胆。当混在其它亚种的种群中时,海角蜜蜂是所谓的“社会性寄生物”( social parasite)。这些工蜂不但“违规”产卵、争抢王台,还散发外激素让普通工蜂将它们当幼蜂哺育,几乎享受着蜂王待遇。它们垄断生殖,致使原种群渐渐崩溃,然后转而入侵新的蜂群。
海角蜂群还能在缺失蜂王的情况下维持数月,直至产生新的蜂王。上世纪90年代,南非每年有50000蜂群因其毁于一旦,海角蜜蜂因此被欧美、澳大利亚等地禁运。海角蜜蜂的社会性寄生现象是在巴西引入南非的蜜蜂后首先发现的,幸亏当时处理迅速,不然可能就不叫海角蜜蜂了。
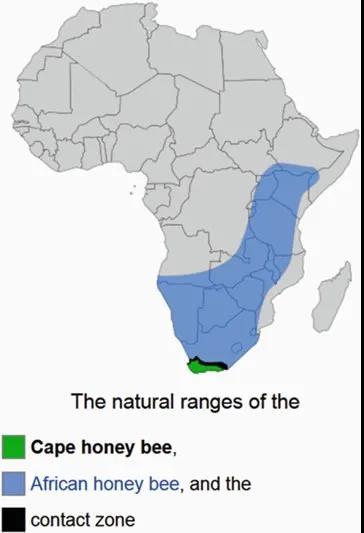
图3. 海角蜜蜂的地理分布十分狭小,仅仅局限在非洲大陆的最南隅(绿色部分)。
大家都知道,蜜蜂的身体是黑黄条纹的,但在海角蜜蜂的腹部,黑色的比重更大(图1),这让我们可以方便地辨识出它。能自己任命领导(蜂后),自产大军的海角蜜蜂,可以算是黑化的打工人了吧。
海角蜜蜂是如何把全套染色体传给后代的呢?
海角蜜蜂的卵也是会经历减数第二次分裂的,但是染色体分离完成之后,并不会马上形成核膜和细胞膜。四对染色体,也就是四个原核(pronuclei)排成一列,然后中间的两个原核(包含的两套染色体来自非姊妹染色单体)发生融合(“中央融合”),恢复为二倍体。海角蜜蜂的工蜂总是进行孤雌生殖,而蜂后总是进行有性生殖。

图4. 一般蜜蜂与海角蜜蜂繁殖模式对比。
孤雌生殖是有弊端的,最突出的弊端便是基因组杂合性的缺失,这会降低物种的适应性。减数第一次分裂中期,染色体会发生重组,然后再经历前面提到的“中央融合”过程。如果某个基因所在的染色体片段与对应的非姊妹染色单体的片段已经交换了一遍,在最终将发育成雌性的卵中,两套染色体上这个片段将是相同的拷贝。对任一个基因位点,杂合度应该丢失1/3,因为在“中央融合”过程中,选定任一个染色单体,剩下的三个里面会有一个是和它一模一样的。
杂合性缺失(loss of heterozygosity)是不利于生存的。一个重要原因是,一些缺陷的隐性基因对应的不利性状会表现出来。比如在癌症中就经常发现有杂合性缺失,在缺失区域常常发现肿瘤抑制基因亦缺失。研究者曾认为,海角蜜蜂群体应该会衰退得很快。但是三十余年过去了,这种情况并没有发生,这或许是因为海角蜜蜂具有社会性寄生的特性。研究者推测,除了csd基因的纯合致死效应,应该还有其它未知的机制在维持海角蜜蜂的基因组杂合度。总之,海角蜜蜂携带着许多谜团,还等待着我们去解开。
最后一个问题,海角蜜蜂这样叛逆的“打工人”是什么时候出现的呢?
研究者发现,所有这些海角蜜蜂都是同一只变异工蜂的后代, 这只工蜂大概生活在1990年。你可能要问,这到底是如何找到的?这里就要提一下生态学的一种重要研究手段——卫星DNA。
卫星DNA是一类高度重复的基因序列。它是一种分子标记,在生态学中常用来代表更长染色体片段。之所以叫“卫星”,是因为在基因组DNA氯化铯密度梯度离心实验中,它总是分布在主条带以外,好像卫星的位置。
卫星DNA有一类称为“微卫星”,由2-4个碱基的序列大量串联重复而成。分析海角蜜蜂样本的大量微卫星位点,科学家发现,除了少数突变,所有个体在这些微卫星位点上,要么是该微卫星序列仅有的两个版本的杂合子,要么是这两个版本其中一个的纯合子。这意味着,它们全部来自一只变异蜜蜂,都在这只“小祖宗”的孤雌生殖谱系上——堪称现实世界的Smith。
这听起来有点惊悚,但你也可以换个角度来理解这个事情。海角蜜蜂变异毕竟是小概率事件,这样的事件发生过一次的可能性当然要比独立地发生两次的可能性大得多(后者概率是前者概率的二次方)。
其实,海角蜜蜂的地理分布十分狭小,仅仅局限在非洲大陆的最南隅(图3)。对于这一区域的养蜂人来说,海角蜜蜂是可爱的存在——好打理,更适应当地的生态环境,更耐受瓦螨(Varroa destructor,是养蜂业的首要害虫),在商业价值上完全可以替代普通蜜蜂。而对研究者来说,海角蜜蜂则是最有意思的一个蜜蜂亚种。
参考文献
[1] Baudry E, Kryger P, Allsopp M et al (2004) Whole-genome scan in thelytokous-laying workers of the cape honeybee (Apis mellifera capensis): central fusion, reduced recombination rates and centromere mapping using half-tetrad analysis. Genetics 167:243–252.
[2] Beye, M., Hasselmann, M., Fondrk, M.K., Page, R.E., Omholt, S.W., 2003. The gene csd is the primary signal for sexual development in the honeybee and encodes an SR-type protein. Cell 114, 419e429.
[3] Jordan Lyndon A, Allsopp Michael H, Oldroyd Benjamin P, Wossler Theresa C and Beekman Madeleine 2008. Cheating honeybee workers produce royal offspring. Proc. R. Soc. B.275345–351
[4] Oldroyd Benjamin P., Yagound Boris, Allsopp Michael H., Holmes Michael J., Buchmann Gabrielle, Zayed Amro and Beekman Madeleine 2021.Adaptive, caste-specific changes to recombination rates in a thelytokous honeybee population. Proc. R. Soc. B.2882021072920210729.
[5] Oldroyd, B., Allsopp, M., Lim, J., & Beekman, M. (2011). A THELYTOKOUS LINEAGE OF SOCIALLY PARASITIC HONEY BEES HAS RETAINED HETEROZYGOSITY DESPITE AT LEAST 10 YEARS OF INBREEDING. Evolution : International Journal of Organic Evolution., 65(3), 860-868.
[6] Rabeling C. Social parasitism. In Encyclopedia of Social Insects (ed. Starr, C.) 836–858 (Springer, Berlin, 2020).
[7]
话题:

0
推荐
财新博客版权声明:财新博客所发布文章及图片之版权属博主本人及/或相关权利人所有,未经博主及/或相关权利人单独授权,任何网站、平面媒体不得予以转载。财新网对相关媒体的网站信息内容转载授权并不包括财新博客的文章及图片。博客文章均为作者个人观点,不代表财新网的立场和观点。
 京公网安备 11010502034662号
京公网安备 11010502034662号 {kind=link}