阅读:0
听报道
每一个科学难题的背后都有一盏点亮的明灯,只不过是这盏明灯被幽闭在不透光的密室里,使得外头的人无从知晓。探索者的努力无非是在厚薄不等的墙上砸开一条裂缝,直到看见里面透出的一线光明。
撰文 | 翁羽翔(中国科学院物理研究所 软物质物理实验室)
1 引 言
2020年6月23日,《中国科学:化学》在线发表了题为“Dynamical and Allosteric Regulation of Photoprotection in Light Harvesting Complex II (高等植物光系统II捕光天线蛋白实现光保护功能的动态及变构调控) ”一文[1],一项马拉松式的研究工作终于暂告一段落。先前和朋友聊起这篇论文探索过程之艰辛、发表过程之坎坷时,朋友极力鼓励我把这段经历写成文字,也许对年轻的研究者有所启迪和借鉴。然而在提笔的时候,脑海中总有一个挥之不去的名字:卞和。《卞和献璧》是一个大家耳熟能详的故事,是中学课文中的一篇文言文,讲述的是卞和献宝的经历。卞和在楚厉王在位的时候,认定自己找到的璞石是一块稀世宝玉,就把它献给厉王。厉王命玉工看后认为只不过是一块普通的石头,卞和由此落个欺君之罪而失去一条腿。来年卞和不服,又来献宝,结果失去了另一条腿。厉王死后文王继位。新君继位,革故鼎新,于是便派人去找卞和,此时的他已在山中哭泣了三日,泪尽继之以血。文王令玉工剖璞验石,这才有传于后世的和氏璧。尽管卞和的执着为后人所称道,但也引来后人的质疑:(1)卞和凭什么坚信他捡到的石头就是宝玉?就凭他看到凤凰曾经栖落在这块石头上?(2)卞和为什么非要去献宝?(3)卞和为什么不自己找玉工先剖析璞石,而非要等君王身边的玉工来评判?虽然卞和最后为自己洗净了冤屈,但结果还是让人觉得有些侥幸。在我看来,与其说卞和只是为了献宝,还不如说是在坚守他自己的信念:凤凰非宝石不栖。尽管现在看来这一信念有点可笑,可仔细想来,我们自己这些现代人身上或多或少也能够折射出卞和的影子。
2 强光下的光合作用——一个少年的偏见
江南的仲夏,田野中墨绿色的水稻在正午的烈日下拔节、扬花、灌浆,直到如金粒般的成熟。辛勤劳作的农人难得有一段清闲的日子,企盼着来日有充足的阳光,好去充孕每一株谷穗。
叶簌知风起,
蛙鸣道雨停。
乐极农闲月,
灯下伴书琴。
这样的日子,分不出是唐是宋,还是元、明、清。直到西学渐近,科学昌明的年代。如果时光能够穿越,我一定要去唤醒那位面带饥色的少年:他经常在仲夏的烈日下冲着田野痴痴地发笑,坚信每一株水稻正在烈日下不停地进行光合作用,而光合作用的名词也是不久前刚从他的老师那儿听来的。烈日普照—光合作用—粮食丰收—碗中的白米饭,一幅多么激动人心的画卷,难怪少年被这正午的烈日陶醉了。
然而,现代科学研究表明:在自然环境中,太阳光的辐照强度可以在短时间内呈现出十几倍的差异。植物不仅要在低光照条件下实现高效捕光,并把激发能传递给反应中心进行后续的光能—化学能转换,还要在高光强条件下将过量激发能以热的形式耗散掉,从而切断能量传递通路,避免过量光能造成的辐射损伤,实现光保护功能。也就是说,在仲夏正午的阳光下,水稻还有其他植物是不进行光合作用的,而是忙着自救,避免过量光能对反应中心带来的损伤!
3 高等植物主要捕光复合物的“中国晶体”——于无声处听惊雷
现代科学前沿领域的思想和实验探索工具越来越复杂,分工也越来越细。顶尖的科研成果很少会出自寒门,大都得益于优良的学术传承和发扬光大。中国科学院植物研究所在植物光合作用生物化学研究领域至少已历经两代人的坚守和努力——汤佩松院士和匡廷云院士。中国科学院生物物理研究所的X射线晶体学研究方向则以人工合成胰岛素的晶体结构解析而扬名于世,这一学派代表人物有梁栋才院士、常文瑞院士、饶子和院士。这两大学派珠联璧合,催生了中国光合膜蛋白原子级分辨率晶体结构解析的学术高地。匡院士作为第一批国家重点基础研究发展规划项目 (973项目,1998—2002年) “光合作用高效光能转化的机理及其在农业中的应用”的首席科学家,对我国的光合作用基础研究起到了组织和推进作用。作为该项目的代表性成果,2004年3月18日, 常文瑞院士和匡廷云院士领导的团队在 Nature 上以主题论文的方式发表了“菠菜主要捕光复合物 (LHCII) 2.72 Å分辨率的晶体结构”的学术论[2],这是国际上第一个用X射线晶体学方法解析的绿色植物捕光复合物高分辨率空间结构。
论文发表之前,笔者有幸参加了在香山双清别墅举办的项目结题交流会。因为参加过几次年度工作交流会,对一些课题的进展有大致的了解。菠菜主要捕光复合物晶体结构解析的工作进展照例由常先生作汇报。先前几次会议对常先生的印象是不苟言笑,作报告有板有眼,喜怒不形于色,当然也给年轻人留下了一种威严的感觉。
和往常汇报的风格一样,常先生展示了 LHCII 的针状晶体和收集到的X射线衍射数据。记得上一次汇报的时候,常先生讲到这,底下的听众一阵激动,当大家期待着常先生给出晶体结构的时候,他非常平静地说道:“晶体在X射线的辐照下,崩溃了!”底下又是一阵惋惜的骚动。
这次常先生讲到这儿的时候,大家都有心理准备,大不了晶体又崩溃了!可这次没有,常先生先不紧不慢地给大家展示了 LHCII 具有和20面体病毒一样空间点群的单晶元胞结构,然后才不温不火地展示了 LHCII 原子级分辨率的晶体结构,当时整个会场被镇住了,足足有大约5分钟的时间会场上鸦雀无声,我是第一个从这种震撼中清醒过来的,便带头鼓起掌来。在持续的掌声中,常先生还是一言不发地站在讲台上,直到匡先生热泪盈眶地拿起了话筒,开始讲述个中的曲折,而我在底下悄悄地按下了快门 (图1) 。会后常先生专门打电话叮嘱我在论文正式发表前,不要外传有关晶体结构的照片。

图1 (左)匡廷云院士和常文瑞院士在2003年召开的973结题汇报会上;(右)2005年匡先生给笔者布置从“中国晶体”到“中国机理”的研究任务
LHCII 原子级分辨率晶体结构的解析是中国科学家的骄傲。匡廷云院士在不同的场合提到 LHCII 的晶体结构被国外同行科学家称为“中国晶体”,以此勉励国内从事物理、化学与生物学交叉研究的学者努力将该蛋白质的工作机理弄清楚,做出与“中国晶体”相称的“中国机理”来。这便是“中国晶体”和“中国机理”说法的由来。对于“中国晶体”和“中国机理”的说法,物理学家戴希教授认为,不宜人为地用狭隘的地域概念去割裂科学,本人深以为然。对于光合作用研究起步较晚的我国,这样的说法更侧重于对后来者的激励吧,现代科学的发展离不开国际间的交流和合作!对于物理所人,冲击“中国机理”便成了一项当仁不让的使命。
4 三人成一众,聚散靠温度——一个令人心动的猜想
LHCII 在自然界主要以三聚体的形式存在,可以简化成三个外切的椭圆 (图2(b)中绿色填充部分)。其中,蓝色三联小球表示镶嵌在蛋白中的叶绿素 b,相应的红色三联小球为叶绿素 a。如果用虚线画出两个圆环分别表示内圈和外圈的相互作用色素分子,可以看出,环上的色素分子排列方式与光合细菌捕光天线中排列成圆环状的细菌叶绿素分子 (图2(a)) 有很大的相似性。对于光合细菌环状捕光天线的量子力学研究表明,细菌叶绿素分子通过偶极相互作用,形成电子态相干的离域态,可以向邻近的捕光天线进行无方向限制的高效传能[3]。
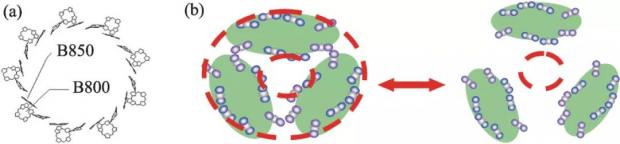
图2 (a)光合细菌捕光天线蛋白—色素复合物(LH2)中细菌叶绿素分子的空间排列示意图;(b)LHCII通过变温诱导解聚/聚集实现高效捕光和光保护功能间切换的猜想示意图
2005年初,在与中国科学院化学研究所张建平研究员的讨论中碰撞出了思想火花,受到光合细菌捕光天线分子结构的启发,高等植物三聚体的两个色素环会不会也有这样的功能?内环色素分子可将能量传递给外环,外环色素分子和相邻的 LHCII 三聚体之间可进行无方向限制的高效传能。如果温度升高,三聚体可能发生解聚,传能环被拆解,从而阻断传能通道,实现光保护功能。等温度降低后三聚体结构又可以恢复,如此周而复始,实现高效捕光与光保护之间的可逆切换(图2)。
图3所示照片是2005年1月22日在中国科学院化学研究所讨论时黑板上的原始记录,左边的粗褐色内切圆代表 LHCII 三聚体的外圈传能环,黄色的外切圆表示内圈传能环。右侧的白色椭圆环 (内标有D1-D2-Cyt559) 表示光系统 II 的反应中心与捕光天线的接触部分。LHCII 通过单体 (M,monomer) 到二聚体 (D,dimer) 及三聚体 (T,trimer) 之间的聚集平衡,“开合自如”地调控了两个传能环,启动了蛋白质分子机器。讨论中提出运用脉冲升温 (T-jump)、核磁共振 (NMR)、单分子光谱及分子生物学等手段来研究这一动态转换过程。
上述想法简单易懂,而且貌似不需要涉及过多的蛋白质动态结构细节,就很自然地把它作为创新点“具有生物活性功能的 LHCII 在类囊体膜中是以三聚体的方式存在的,我们拟将 LHCII 三聚体的动态聚合/解聚合作为研究 LHCII 通过蛋白质动态结构变化实现高效捕光功能和有效光保护功能之间切换的突破点”去申请国家项目,但是一直没有申请成功。虽然这个激动人心的猜想到2017年才彻底破灭,但却激励了10余年间不懈的探索和研究条件的积累。

图3 2005年翁羽翔、张建平讨论后黑板板书照片(讨论纪要为张建平整理)
5 CO中红外激光器——有钱的玩钱,没钱的玩命
1999年回到中国科学院物理研究所工作,当时尚未申请到百人计划的支持。组里最好的设备是先前三个组合买的一台进口YAG脉冲激光器,大家轮流使用。角落里还有一台用紫色天鹅绒布覆盖着的、由石英杜瓦瓶液氮浸式冷却放电管构成的中红外一氧化碳 (CO) 分子气体激光器。这是傅克坚研究员 (傅作义先生女儿) 的课题组留下的[4],闲置在那里应该有5—6年之久,了解这台激光器的人都已经离开了这个实验室。当时在思考如何利用现有的条件开展有意义的研究,而不至于虚度光阴。碰巧在海淀图书城买到一本由国内多位著名科学家撰写的《21世纪的100个科学难题》[5],其中有一篇是中国科学院生物物理研究所王志珍院士 (后来有幸与其合作) 写的文章“中心法则的空白——从新生肽到蛋白质”,谈到“蛋白质折叠问题”是科学难题,联想到我在 Emory 大学做博后时认识的乔治亚理工学院 (Georgia Institute of Technology) Mostafa El-sayed 教授的博士生王建平,他当时正在开展利用脉冲升温—纳秒时间分辨中红外光谱研究蛋白质折叠动力学,于是就下决心开展这个方向的研究。
第一件事是如何让CO激光器运转起来,在找不到知情者咨询的情况下,想到去所科技档案室碰碰运气,看看有没有相关的资料存档。没想到,李春芳老师居然从档案室给了我一个厚厚的资料袋,里面不但有研制任务书,还有图纸及仪器验收指标,关键还有这台设备的研制单位大连理工大学的联系人和电话。凭借这个电话号码,找到了大连理工大学的于清旭教授,从此开展了后来持续多年的CO激光器升级换代工作。2001年开始连续三年向自然科学基金委员会递交应用脉冲升温—时间分辨中红外激光光谱研究蛋白质动态结构的申请,直到第三次申请才终于获得了面上基金的资助。多年后才知道,项目进入会评阶段多亏了当时只有一面之缘的大连化学物理研究所李灿院士的支持。期间在于老师的指导下,学会了CO激光器的出光调试,但是这台老激光器一旦连续运行起来,问题便层出不穷,不是变压器被烧坏,就是大电容被击穿,而且激光稳定性差,就连真空橡胶抽气管用几回也会穿孔破裂,如此等等,不一而足。记得那年冬天,几乎十个指头都缠着胶布。有一次无意中看到一篇资料讲到臭氧对人体皮肤的损害,联想CO激光器放电后石英管中会残存大量蓝色的液态臭氧,不仅腐蚀人的皮肤,肯定也会腐蚀真空橡胶管,由此一举解决了手指皲裂和真空橡胶管被腐蚀的问题。那段时间正好我父亲来北京,很少有时间陪他出去转。老人家从乡下来,满怀希望地想见识一下在中国科学院物理研究所就职、多年游学海外的“海归学子”。看到我每天早出晚归,忙忙碌碌的样子,父亲很是失望,在我家住了一个月就回去了。临走时扔下一句话:“都以为你是个海归,我看你就是一个打工仔”。是的,对不起了父亲,我本来就是一个打工仔,不同的是,这回是为自己打工,为自己的国家打工。
还有一个问题是光路的实时调节。红外激光看不见,因此设计激光器时在镀金的端面反射镜上开了一个 5 mm 的小孔,引入氦氖激光作为指示光。激光放电管由中间的正极 (零电位) 和石英管两端各一个负极构成。正负电极之间的电压为7千多伏。接线柱在激光器靠墙的一侧,但裸露在空气中,平时不会碰到接线柱。在调节谐振腔光路时,需要两人配合,一人的手臂要从靠墙的一面空隙中伸到输出镜侧的定位光阑处,通过开闭光阑查看指示光的位置,指挥另一人调节端面镜的反射光路。实际操作中,我担当“指挥员”的角色。为了安全起见,先在地上铺上绝缘的胶垫,再穿上胶底拖鞋,心想即使碰到负极,身体也只不过是个等电势体,应该不会发生什么意外。在一次调光路过程中,我和张庆利同学(该方向第一个研究生)都穿着短袖。我左手臂无意中碰到电极,而恰在这个时候,张庆利的手臂碰到了我的右胳膊,顿时感觉到一股电流涌向心脏,引起心脏疯狂地跳动,令人窒息。而这时左手腕带的金属表和皮肤之间也放起电来,除了蓝色的电光外,还发出一股焦糊味。奇怪的是,张庆利居然一点感觉也没有,原来是 7600 V 电压通过他 (零电位) 把我给接地了!2006年在厦门大学召开的全国跨学科蛋白质大会上,我在报告中亮出了这句感悟:做科研是“有钱的玩钱,没钱的玩命”,没想到这成了那届会议被引用最多的一句话。
6 毕业就业的压力——一篇文章一条命
脉冲升温—纳秒时间分辨红外光谱研究蛋白质动态结构的原理是,红外光谱能够识别蛋白质的二级结构,如α-螺旋、β-折叠、无规卷曲、环折结构及分子间氢键等。而激光脉冲升温方法可以给蛋白质一个快速的扰动,一般由脉宽为10 ns的脉冲激光激发水或氘代水的泛频吸收,可在20 ns时间内引起体系约20 ℃的温升,比传统的反应停留法测定蛋白质折叠动力学的时间分辨率高出几个数量级。然而常规的掺钕YAG激光器1.064 μm的基频输出无法直接用作加热脉冲,必须经过一个以高压氢气为介质的拉曼频移器,将基频红移到2 μm左右。拉曼频移器看起来很简单,在一段不锈钢管的两端分别安装上石英窗口。但要保证其在几十个大气压下安全工作,谁也没有把握。2002年访问日本东京大学的一个光谱实验室,听说一个充有80个大气压的拉曼频移器出了事故,石英窗片被高压气体推出,在对面的墙上砸出了一个大洞,幸亏没有伤着人。尽管有风险,还是想由自己设计加工,并就此事请教许祖彦院士。许先生告诉我们,清华大学的娄彩云教授做过拉曼频移器用以产生通讯波段的激光 (1.54 μm,用甲烷做介质)。就这样从娄老师那里拿到了拉曼频移器,额定安全工作气压为20个大气压。实验中激光转换效率太低,王莉同学 (北京理工大学联合培养的研究生) 把气体池加长,又在娄老师的指导下提心吊胆地把气压一点点提高到30个大气压,总算满足了实验要求。这时张庆利同学也把CO稳定性提高到了较为满意的水平,与加热脉冲激光联用后,实现了探测光吸收变化率 (ΔOD) 测量精度为10-3的基本要求[6]。当时经费缺乏,连王莉的工资都开不出来了,于是硬着头皮去找杨国桢院士帮我想想办法,以解燃眉之急。没想到杨先生听完我说的困难后,爽快地把他当年的院士个人支配的创新经费(3万元)转给了我。
2006年叶满萍同学开始了LHCII三聚体的变温红外和脉冲—升温时间分辨红外光谱研究,样品通过匡先生介绍由中国科学院植物研究所王可宾老师提供。当时面临的最大问题是单波长扫描测量的瞬态红外光谱重复性不好,于是将红外光分成两路,一路探测样品,一路作为参考光,实时扣除CO激光强度抖动,尽管数据质量明显提高,但重现性还是差强人意。最后只好从几十套瞬态光谱数据中挑出几套自认为可靠性高的进行平均,但仍然难以确定光谱测量的准确性。2008年5月,叶满萍毕业并联系好去东京大学做博士后,按计划9月份去日本。看着这一堆尚未有头绪的数据,就和她商量,能否再利用去日本前这段空档时间,多重复几套数据,给出一个可信的结果?得到的答复是:“从6月6号开始,我已经不是物理所的人了!”,言外之意是老师的指挥棒失灵了。尽管回答让我感到有些突然,冷静下来还是深刻反思一下为什么会这样?连实验技能已经很熟练的学生也在刻意回避和逃离这个课题,换个新手会有改善吗?答案是否定的,仔细研究实验数据后,得出的结论是,测量精度必须提高一个数量级,即达到10- 4 ΔOD 才能够获得可信的数据!没有别的捷径,只能改造升级实验设备!而且必须抛弃使用拉曼频移器这一通用做法。因为实验发现由于高能量激光对气体池的加热效应,导致拉曼频移输出光在空间位置的重现性差,进而导致加热激光和探测光焦点在样品中的空间重合无法保障。另外CO激光输出功率的稳定性也有待于进一步提高。新的方案是将加热脉冲改用掺钬的YAG激光,直接获得2 μm的基频输出。但是这种激光器没有商用产品,要自己研发。
2008年10月获得了中科院重点仪器研发项目的资助,其中研制掺钬YAG激光器的工作由中国科学院安徽光学精密机械研究所的合作者承担,新一代脉冲升温—纳秒时间分辨中红外瞬态吸收光谱仪的调试工作由李得勇同学负责。记得第一次在新的设备上看到瞬态信号时,原来的热切希望被活生生地浇灭了:仪器的电磁干扰信号居然高达1.5 V,主要来自于钬激光电源。我把多年积累的经验用上后,总算把噪声降到了10 mV的水平,然后和李得勇讲,10 mV的噪声水平是我做博士年代的水平,20多年过去了,技术方法总归有很大的长进吧,下面就看你的了,我的要求是毫伏量级。经过各种试错(trial-and-error),终于实现了这个目标[7]。
2015年中国科学院物理研究所科研简报“高精度脉冲升温—纳秒时间分辨中红外瞬态光谱仪的成功研制”写到:2015年5月27日,Review of Scientific Instruments发表了中国科学院物理研究所软物质院重点实验室翁羽翔研究组的一篇题为 “A Q-switched Ho: YAG laser assisted nanosecond time resolved T-jump transient mid-IR absorbance spectroscopy with high sensitivity”的仪器研制论文[8],报道了新一代具有国际先进水平的脉冲升温—纳秒时间分辨中红外吸收差光谱仪的成功研制;包括研制高稳定连续输出可调谐一氧化碳中红外激光探测光源,以及研制新型的脉冲激光加热光源,即空间模式稳定、输出能量稳定的纳秒调Q 的Ho: YAG 脉冲近红外激光光源 (2.1 μm,与中国科学院安徽光学精密机械研究所吴先友研究员合作) 。该设备对蛋白质细胞色素c的脉冲升温—时间分辨中红外光谱测量结果表明,在蛋白质酰胺 I'光谱范围 (1600-1700 cm-1) 内达到的平均测量精度为2×10-4 ΔOD。简报的开头是这样写的:“十年磨一剑,不敢试锋芒,再磨十年剑,泰山石敢挡”。现在看来LHCII就是一块用来试剑的泰山石。

图4 作者(左)和李姗姗(右)向王志珍院士(中)讨教蛋白质折叠动力学问题(2010年12月摄于生物物理所王先生办公室)
和李得勇同学同年入所的李姗姗同学则投入了LHCII的研究工作中。由于研究工作一直没有获得实质性的结果,也不好意思再向植物所要样品了,最好的解决方案是自己能够提取样品,并由此掌握提取蛋白样品过程中所添加的试剂成份和含量。幸运的是,中国石油大学黄岛校区黄方教授组里的刘双老师愿意教我们,李姗姗就在那里学会了从菠菜叶片中提取LHCII的实验方法。由于仪器研制过程中碰到许多预想不到的困难,李姗姗没有等到新的设备,而LHCII稳态光谱的数据尚不能够说明问题,她离毕业要求还差一篇论文,就提出把LHCII的初步结果整理后投给国内的期刊,我没有答应。在延期毕业的日子里,李姗姗只好应用旧的脉冲升温—时间分辨中红外光谱仪研究北京大学生命科学学院昌增益教授提供的热休克蛋白,在修改稿返回给Scientific Report 编辑部漫长的三周时间等待中,延期毕业、就业竞争以及生活与家庭,似乎所有的成功与否都维系在一篇论文是否被接收上,来自数方面的压力差一点将她压垮了。差不多是第三周的一个晚上,晚饭后照例查一查邮件,当看到了编辑部的接收函时,便一下子从椅子上跳了起来,大声嚷嚷“接收啦!接收啦!”,以致我太太跑过来看到我当时的情形,冷冷地说了一句“你们这些搞科研的,一个个像范进中举似的,不就一篇文章,至于吗?”,我当时的回答是:“你知道吗?一篇文章一条命!”
7 结识高加力教授——无心插柳的机缘
美国明尼苏达大学化学系和超算中心的高加力教授是分子动力学模拟研究的专家,是分子动力学模拟的鼻祖、诺贝尔化学奖得主哈佛大学Martin Karplus教授的高足。我和他在2015年基金委物理化学处重点项目答辩会上初次见面,高老师来做申请答辩,他与吉林大学理论化学研究所开展合作研究工作,我是当时的评审专家。那年评审会印象深刻,年轻学者如复旦大学的刘志攀教授等胜出,而两员老将高老师还有中国科学技术大学的严以京教授则铩羽而归。第二年高老师继续参加答辩,项目是应用分子动力学模拟的方法计算蛋白质二级结构的红外光谱,精度达到 1 cm-1。这样的申请报告对我而言不啻是高山流水之音。一激动,就做了一番即席发言。高老师那年终于拿到了项目,也许就是因为这一番发言,他便知道了国内有位运用红外光谱解析蛋白质二级结构的翁羽翔。那年9月份,高老师和他的同事李辉教授突然造访中国科学院物理研究所,就在物理所“总理咖啡屋”一叙。寒暄过后,就切入正题,我非常希望能够和这样的高人开展合作,于是就在咖啡屋的电子屏幕上给高老师介绍起了高等植物捕光天线的非光化学淬灭和我们的LHCII三聚体动态聚合/解聚调控的想法和研究进展。
在长期的进化中,植物发展出多种不同时间尺度与空间尺度的光保护机制,其中一种极为重要的机制叫做非光化学淬灭。非光化学淬灭发生的场所主要在类囊体膜,其淬灭位点包括光系统II主要捕光天线三聚体(LHCII trimer)与内周辅助捕光天线单体 (CP29、CP26、CP24)。捕光天线将过量的激发能在微秒时间尺度内以热的形式耗散到环境中。尤其是捕光天线 LHCII,叶绿体中的含量最高约占60%左右。晶体结构表明,LHCII单体包含6个叶绿素b,8个叶绿素a,与4个类胡萝卜素分子:两个叶黄素 (lutein),一个新叶黄素分子 (9'-cis Neaxanthin) 以及花药黄素 (antheraxanthin,叶黄素循环的中间体) 。其中,LHCII 的蛋白质骨架由三段跨膜α-螺旋(A, B,C)构成,A和B呈剪叉式排列,交叉点由两对带正负电荷的氨基酸 (Arg70-Glu180,Glu65-Arg185) 通过形成盐桥锁定;类囊体腔侧包含一段双亲性α-螺旋D与另一段酸碱两性的松弛螺旋310-螺旋E;螺旋E和C之间由一段反平行β股结构相连。叶绿素分子“镶嵌”在蛋白质骨架上,彼此之间以特定的取向和距离有序分布,在捕获光子与传递光能的过程中发挥着重要的作用。类胡萝卜素分子主要分布在叶绿素分子的周围,通过及时清除自由基氧和过多的光能,保护植物免受过多光能的损伤,蛋白质骨架为这些色素分子提供结合位点。
当时我们以为已经获得了LHCII三聚体解聚的初步证据,实验方案是这样的:依据晶体结构,LHCII三聚体中,单体界面处存在一些结合的水分子,在室温条件下,没有暴露出来的水分子不会和溶剂氘代水进行氢氘交换,如果发生热解聚,随着温度的升高,结合的水分子会释放出来进行氢氘交换后形成HOD,并且在红外光谱1450 cm-1处出现有别于H2O和D2O的特征峰。前期叶满萍、李姗姗和后来的李昊同学,他们的变温红外光谱的数据都证实1450 cm-1的吸收强度随温度升高而增强。最后希望高老师能够和我们合作,用分子力学模拟揭示LHCII随着温度升高的解聚过程,证实LHCII三聚体动态聚合/解聚的想法,因为到目前为止,稳态和瞬态光谱数据既无法证实,也没有否定该想法。
8 痛苦不一定来自外部的打击,但一定是内心希望的破灭
大约半年后,高加力教授第二次来到物理所,同样在咖啡屋,只是演讲的主角换成了高老师。高老师先展示了由24万多个原子构成的LHCII三聚体、磷脂双层膜和水分子构成的起始模型,然后演示了三聚体在高温下存在的一个开合运动,以及界面结合处水分子随温度升高逐渐脱落的过程。当看到水分子的确随着温度的升高从界面处脱落,这一点和实验相符时,感到很高兴。但接下来是令人困惑的问题:分子动力学模拟根本没有揭示三聚体存在随温度升高而解聚的迹象!结合植物所卢从明研究组2007年在Plant Physiology 上发表的实验工作[9],他们的结果表明LHCII三聚体随着温度的升高确实发生了非光化学淬灭 (NPQ,non-photochemical quenching),但没有发生三聚体的解聚,而是热诱导三聚体聚集过程。面对上述事实,不得不放下幻想,眼前一片迷茫,就像吹灭了十多年来照亮在黑暗中摸索的一盏油灯,此刻又重新陷入了彻底的黑暗。怎么办?投入了这么多的人力、物力和时间,难道就此打住?冷静下来后意识到原来为什么这么执着于这个想法,有深层次的原因,那就是对海量生物文献的恐惧,因为只要物理模型成功,就没有必要去了解生物学上的细节。在当前这种进退维谷的情形下,唯有扎扎实实地做好文献调研,理清学术脉络才有重新启航的希望。接下来在查阅大量与荧光非光化学淬灭的相关文献后认识到 (到论文发表为止,阅读了近500篇论文) ,国际上对于NPQ的机制原来一直是有争论的,尽管如此,以英国伦敦玛丽皇后大学的Alexander Ruban教授为代表的一派提出的机理十分吸引人,也基本上处于主流地位,即一对交叉排列的叶黄素分子,各带一个叶绿素a分子,形成两对叶黄素—叶绿素色素对,当色素对中叶绿素与叶黄素分子间距较远时,叶黄素分子将光谱蓝区捕获的能量传给激发态能量较低的叶绿素分子,实现高效捕光;如果两者靠得很近,则叶绿素将激发态能量传给叶黄素的光学暗态,再由该暗态将激发能以热的形式耗散掉,并把调控过程归结于蛋白质的构象变化。然而蛋白质构象是如何实现这一“智能”调控的,仍然是一片空白。于是对LHCII变温红外光谱进行仔细的解析,找出各个红外吸收峰在变温过程中的相关性,结合晶体结构信息,尽量找出空间相邻的二级结构的对应关系,希望在蛋白质构象变化方面有所进展。最重要的突破是指认了310螺旋E,由于其结构比常规α-螺旋更为松弛,出现在α-螺旋(1653cm-1)的蓝区(1658 cm-1)。2017年4月,在对蛋白质构象变化有一个大致思路的基础上,给李昊同学布置了一个任务,立即停下他手头在做的与白内障相关的α-晶状体项目,利用新建的设备,开展LHCII脉冲升温时间分辨中红外光谱研究,当时估计实验也就需要三个月的时间,没想到断断续续持续了三年。
继2008年之后,终于又能够应用升级改造后的设备再次回到叶满萍同学中断的地方。因此对于这套数据尤为期待,恰好叶满萍这段时间也在组里做访问学者,无意中见证了激动人心的时刻。李昊对实验数据质量的追求有一种“洁癖”,实验细节把控极严。我在比较了几届学生的LHCII红外光谱后发现,他做的数据质量最高,特别是氘代水溶剂中来自水的吸收干扰,他排除的最好。两片光学窗片夹一层50 μm 厚的含表面活性剂的LHCII液体样品,他在手套箱中装样,做的透明均匀,不含气泡。记得这一貌似简单的装样过程,先前有同学就是无法消除样品池中的气泡,连续做一个多月,哭着来找我,最后我也没有想出好办法,只能靠多装样,凭手气。也正因为这种秉性,看他的数据要催得勤快点。李昊终于告诉我T-jump数据做好了,我在办公室等了半天也没收到邮件,忍不住跑到学生办公室去找李昊。李昊没在办公室,我冲着叶满萍喊了一嗓子 “李昊在哪,他把数据藏着掖着干嘛?!”“他刚才还在,数据在电脑里”,于是叶满萍伸手去碰了一下李昊电脑的鼠标,唤醒了休眠中的屏幕,跳出一幅瞬态光谱图,叶满萍大叫一声:“翁老师,你看李昊的光谱不是和我的一模一样吗!”我压抑住心头的狂喜,冷静地说,你再比较一下你们俩的动力学数据的差异,这回叶满萍服了:李昊的动力学给出了十分平滑的蛋白质去折叠过程的曲线,而满萍的动力学曲线则含有很大的振荡干扰,高下立判。磨刀做设备的功夫终于在这一刻收到了回报。
9 全原子分子动力学模拟——蛋白质动态结构的虚拟实验
科研界做实验的往往更相信自己的实验结果,而把理论计算及分子动力学模拟等作为佐证实验结果的锦上添花之作,我也深染这种习气。
高加力教授和王英杰博士早早地获得了分子动力学模拟的结果,而我们却只分析了与界面水分子相关的结果,仅仅用到了海量数据的冰山一角。T-jump实验数据中,我们发现β-折叠、310螺旋的消失过程在动力学上很好地对应了α-螺旋的形成过程,因此请王英杰看看模拟的结果是不是这样。很快王英杰发来了图片,不仅证实上述过程的发生,还问我们实验上是否看到无规卷曲结构也变成了α-螺旋,因为模拟显示,两个单体结合处一个单体的310螺旋E诱导另一个单体α-螺旋D端的无规卷曲变成了α-螺旋。我们在实验上看到无规卷曲结构的消失过程,但从来不敢做这样的结论,因为通常而言只有α-螺旋失稳变成无规卷曲结构,而不是相反的过程。实验中分析出新生的α-螺旋有一个微秒尺度的插膜过程,模拟结果得到证实,实验中还发现谷氨酸羧基的红外吸收强度随温度的变化和荧光淬灭曲线几乎一致,推测囊腔侧的谷氨酸有可能是导致荧光淬灭的触发开关,模拟结果证实一个特殊点位的谷氨酸E94,利用氢键起到了开关切换的助力作用。这样实验和模拟结果相互印证的例子还可以举出很多。全原子模拟中有24万多个原子,随着时间在变化,产生海量的数据,如果没有实验上的线索或者理论的预期,这些细节一不知道从何看起,二不知道结构微妙的变化所蕴含的物理意义。
简单地总结一下,全原子分子动力学模拟结果表明,在升温过程中,蛋白质部分结构单元会发生失水作用,导致结构单元间的疏水作用增强,引起类囊体腔侧一段处于松弛状态的310螺旋E和相邻的 LHCII单体中的一段无规卷曲结构协同转变为α-螺旋,并引起α-螺旋E和D向疏水核内部插入运动,从而在原子尺度上阐释了脉冲升温—时间分辨光谱观察到的蛋白质二级结构的变化。新生的α-螺旋E和D插膜运动引起的别构效应驱动一对交叉排列的跨膜螺旋A和B发生剪切运动,导致附着于跨膜螺旋对上的叶绿素分子与相邻的叶黄素分子之间的间距随温度的升高或酸度的增加而变小。进一步通过量子化学计算,揭示叶黄素分子 (LUT) 暗态 (S1态) 和叶绿素a激发态电子态间的耦合强度在一定的温度范围内,也随温度升高而增强,导致叶绿素激发态的荧光淬灭,并且荧光淬灭主要发生在LUT-Chl612这一色素对上。
这次和高加力课题组实验和理论的配合,彻底改变了我对分子动力学模拟的成见,可以说分子动力学模拟已经达到了蛋白质动态结构虚拟实验的水平,当然人的因素永远是第一位的。
10 热、酸触发的LHCII剪叉式升降运动实现高效捕光到能量淬灭的切换
我们的研究结果表明,LHCII恰如剪叉式升降机一样的分子开关 (图5) ,在热效应或酸诱导的疏水作用下,触发螺旋E与D向疏水内核的插膜运动,将动力分别传给剪叉的两臂螺旋A与B,引起像剪叉式升降机一样的运动,迫使叶黄素分子和相邻的叶绿素分子相互接近,从而实现叶绿素激发态的能量淬灭。从机械的角度,还能够找到剪叉式升降机所对应的关键部件。类囊体腔侧的螺旋E与D为一对顶杠,螺旋D端的残基L206与螺旋B末端残基V80、L84,螺旋E端的残基W97与螺旋A的末端残基F194、F195分别构成了类囊体腔侧的两个顶杠作用力支点。一对交叉排列的螺旋A和B为升降的两个剪切臂,其交叉点处的盐桥 (Arg70-Glu180,Glu65-Arg185) 为铰点。温度升高或酸度增加时,螺旋E和D分别从跨膜螺旋A与B交叉面的内侧将螺旋剪叉臂顶开,导致剪叉角度增加,带动两个剪叉式排列的叶黄素分子做相应的开叉运动,缩短了叶黄素分子与相邻的叶绿素分子之间的距离,增强了色素之间的电子态耦合作用,提高了叶绿素激发态荧光淬灭的效率。尤其在低温条件下,位于腔侧的谷氨酸 (E94) 与螺旋E近端的赖氨酸K99形成氢键,随着温度的升高,螺旋E逐渐朝中心靠近,E94-K99之间的相互作用减弱,而E94与处于螺旋E 中段的谷酰胺Q103形成新的氢键,这说明E94协同参与了螺旋E的插膜过程。上述模型与已报道的分子生物学中单点突变的实验结论相符合,如将E94突变成电中性甘氨酸,会导致非光学淬灭效率的降低。可见E94恰如开关中的触臂,K99及Q103分别为两个触点,当E94与K99相触的时候,系统处于高效捕光态,而当E94与Q103相触的时候,则处于光保护状态。
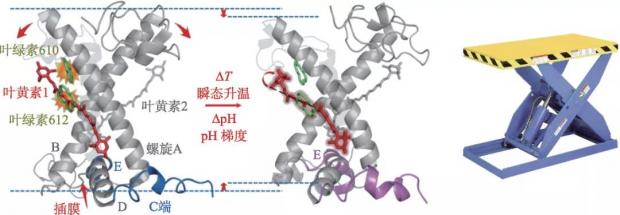
图5 热及酸度驱动LHCII三聚体分子开关剪叉式运动工作原理(左)及剪叉式升降机器(右)类比示意图
进一步从分子机器的角度可得出以下三个推论:
(1)插膜运动速率是能量淬灭过程的速率决定步骤,时间尺度为1.24 μs,因此光合系统能量耗散也应该在这个时间尺度。文献表明,利用光声光谱测定的活体叶片能量耗散时间为1.4 μs;
(2)从机械的角度考虑,囊腔侧螺旋插膜作用需要经过几道环节的传递才能够转变为跨膜螺旋的剪叉升降运动,而传递过程中涉及的机械单元是典型的软物质,因而光合膜刚度越大,力的传递效率越高,能量耗散的效果也就越好,这点也被文献报道的实验所证实。
(3)能量淬灭态对应的是交叉跨膜螺旋对顶角更为张开的状态,对应的光合膜变薄,这一点也与已报道的实验观测相符。
上述分子机制不仅与迄今为止的许多生物实验观测相符,还揭示了植物是如何应用稳态温度和瞬态温度来实现高效能量耗散、同时又可以避免蛋白质热变性。一般情况下,当环境温度为25 ℃时,叶片的温度可达40 ℃。叶片对激发能的耗散弛豫时间为10 μs量级,进一步估算10 μs内吸收的光子数全部变成热量可以引起LHCII三体瞬态温度上升14 ℃,并且该瞬态温度持续时间远小于100 μs的蛋白质变性条件,因此不会引入额外的蛋白质变性,处于能量耗散光保护状态的瞬态温度达到54 ℃,正好处于能量淬灭的最佳温度附近,而蛋白质的稳定性则由稳态温度40 ℃决定。可见自然界利用高瞬态温度实现最优化能量淬灭,同时维持较低的叶片稳态温度以保证蛋白质结构的完全可逆变化。
11 投稿——锤铁成钢的过程
一项经得起时间检验的研究工作的出炉,不但要感谢一路上相扶持的盟友,更要感谢那些曾经出手猛烈捶打的对手。原创性的工作通常会给探索者带来按捺不住的惊喜,恨不得抓住每一个你认识或不认识的人,和他们倾诉你的发现,让他们和你一起分享发现的喜悦。然而科学发现的分享和发表自有它的一套规则,即同行评审制度。出色的工作要经得住无比挑剔的审稿人的捶打,捶打的越猛,就如打铁一样,越能够剔除杂质。然而该项制度设计理想的一面是这样的,不理想的一面和其他行业一样,折射出的是复杂的人性。当作者把论文投给影响因子高一些的杂志时,第一关往往是两周时间,内容和结果先要通过编辑部的预审,过了这一关才能到审稿专家手上。因此,大多数作者都会有诗中所描摹的新媳妇将要拜见公公婆婆的忐忑之心。不同的是,对于匿名审稿,多数情况下不知道审稿人是谁,以致习惯性地从评审意见的语气、用词和眼界去判断审稿人的身份,论文中哪些观点招惹了审稿人,甚至能够读出审稿人当时的心态,这种有点像狐狸和猎人之间斗智斗勇的过程对于初入此道的学生往往会感到迷茫,“翁老师,我们是做科学研究的,难道还要去分析审稿人的心情吗?”不止一个学生向我提出这样的困惑,而我的回答是“科学研究本身是客观的,但科学是人做的,是由人来评判的”。
近试上张水部
朱庆馀
洞房昨夜停红烛,待晓堂前拜舅姑。
妆罢低声问夫婿,画眉深浅入时无。
2017年12月底,收到第一次审稿意见。第一个审稿人的评价是“探索一个十分重要但很少被研究过的问题……总体来说这项工作给人印象深刻,大多时候表述得很好,反映了该领域的新进展”。第二审稿人在对工作给予一个简洁的总结和建议之后,按照编辑部对稿件各方面的标准,逐一进行了回答,同时也列出了相应的问题,非常详细和专业,说实在审稿人提出的一些生物学上的细节我们之前是没有注意到的。按照以往的经验,很快判断出第一个审稿人是这领域的大家,也是我们学术思想的同盟军。根据第二个审稿人如此多的细节问题,以及没有读出一种高屋建瓴的气势,我们判断或许是某位大家手下的博士后。尽管意见很多,综合起来还是很乐观。
对于审稿人意见,尤其是貌似不友好的,习惯上都会放上一周等情绪平静后再来比较客观地逐一琢磨。这次也不例外,当读到“ (叶绿素和β-胡萝卜素分子间) 激发态能量转移效率最高处发生在多烯的骨架中心位置,而不是链端的六元环”时不禁出了一身冷汗:一个生物学的博士后绝对不可能有这样的学识!一定是该领域的大家,而且猜测是对能量淬灭分子机制持反对观点的一位欧洲科学院院士,这也预示着一场“恶战”不可避免。
红外光谱指认蛋白质笼统的二级结构一般不会招来质疑,如果要精确到空间的某一片段,往往需要其他手段的支持。蛋白质氨基酸序列单点突变技术是十分有效的方法,通过改变光谱指认片段的多肽序列,或者在结构上删除该片段,如果对应的红外光谱发生变化或消失,则为光谱指认提供了可靠的支持。我们先前和王志珍院士[10]、植物所种康院士[11]及昌增益教授[12]合作发表的工作都是以这样的方式回答审稿人疑问的。这次审稿人同样刺中了我们的软肋:关键二级结构310螺旋E的红外光谱指认。光合膜蛋白单突变体无法从菠菜中提取,只能通过人工组装的方法来实现。了解到国内只有植物所杨春虹研究员课题组能做这样的工作,而在这个节骨眼上,她到了退休年龄,课题组面临解散,也是爱莫能助。突然想到9年前李恒帮她测过一个突变体的红外光谱,数据好像一直没派上用途。赶忙从学生科研档案中找到了原始数据,在多套原始数据中获得了达到精度要求的红外光谱数据,圆满地回答了这一问题。
在随后第二、第三次回答中,双方争辩更加激烈,而第一个审稿人从此保持静默,不给任何一句评价。在第三次给编辑的回信中,对于第二个审稿人的意见“基于上述通常的做法,我的结论是分子动力学模拟必须给出参与被研究调控过程的关键残基或者结构域,然后对该残基或者结构域进行突变来中断调控过程,从而证实提出的假说”,高老师的回答也是充满火药味:“氨基酸残基突变对某些问题固然有效,然而作为美国化学会旗下的期刊 Journal of Chemical Theroy and Computation 的编辑,可以肯定地说,这不是分子动力学模拟学术圈的标准,科学上对我们完全是不公正的”。这样的交锋结果可想而知,不仅拒稿,主编还直接告诉我们对该工作不再感兴趣了。只好再改投另外的杂志,我们的工作投过去不久,国外的一个工作跟踪而至,利用基因敲除的方法,在一个结构类似的辅助捕光天线膜蛋白中证实了我们工作中理论预测的荧光淬灭色素对。结果是他们的工作发表了,我们的工作又被拒了,当然也收到了针对实验几乎点到死穴的审稿意见,即蛋白质变性温度和荧光淬灭温度问题,如果淬灭温度高于变性温度,则研究结果就偏离了实际情况,可以说是一剑封喉。经过几个月的努力,终于从实验和理论上解决了上述问题。回头一看,国外在这一时期发出了十多篇论文,有的工作为了抢到优先权,从投稿到发表只用3天时间。从发表的论文中发现,原先审稿人拒绝我们的理由已经不复存在,我们把相关论文和申述意见寄给编辑部,主编居然受理了我们的申述,重新审稿。2020年春节,一个难忘的日子,收到了第四次也是最后一次该杂志的拒稿信。尽管如此,还是对主编充满感激之情。经过这几轮捶打,终于明白一个道理,这个工作是难以在国外的高影响杂志上发表的,最后决定投给《中国科学化学》。编辑部邀请了4位审稿人,又提出了一些很好的建议,修改稿终于于5月9日接收。在经历了六轮的投稿,回答了上百个问题后,自以为到目前为止做的最为重要的一项工作,如一颗随风飘浮的种子,终于落在自家的花园里,衷心地祝愿我国自己的杂志越办越好,打造一块引吭高歌的发声地。当把接收消息告诉高加力教授后,他给大家的回信是“这必将成为他们的领域的经典之作”,我把“他们的”改成了“我们的”。6月4号物理所学位评定委员会全票通过,授予在组里工作和学习了8年之久的李昊同学博士学位。
12 结束语
对于科学探索,思想是自由的,而行动则会受到各种条件和规则的牵制,而从事自由探索的这个群体,宛如一群戴着脚链的舞者,在有限的空间中展现出无限的美丽。记得张殿琳院士有一次在物理所春季年度学术交流大会上就学术考核问题,引用了白居易的诗句:
赠君一法决狐疑,不用钻龟与祝蓍。
试玉要烧三日满,辨材须待七年期。
周公恐惧流言日,王莽谦恭未篡时。
向使当初身便死,一生真伪复谁知?
张先生重点强调了“三日满” 和“七年期”。对于崎岖路上的探索者,也许这也是一种回答 (摘自汪国真《热爱生命》):
我不去想是否能够成功
既然选择了远方
便只顾风雨兼程
然而现实是,只有成功,才有话语权。“publish or perish (要么发表,要么走人) ”流传已不止一天了,天下苦秦久已!
参考文献
[1] Li H,Wang Y,Ye M et al. Sci. China Chem.,2020,63:8
[2] Liu Z,Yan H,Wang K et al. Nature,2004,428:287
[3] 翁羽翔. 物理,2007,36:817[
4] 林金谷,王文华,徐欣渡 等. 仪器仪表学报,1991,12:232
[5] 21世纪100个科学难题编写组. 21世纪100个科学难题. 长春:吉林人民出版社,1998
[6] Zhang Q L,Wang L,Weng Y X et al.Chin. Phys.,2005,14:2484
[7] 翁羽翔,陈海龙 等. 超快激光光谱原理与技术基础. 北京:化学工业出版社,2013
[8] Li D,Li Y,Li H et al. Rev. Sci. Instr.,2015,86:053105
[9] Tang Y,Wen X,Lu Q et al. Plant Physiol.,2007,143:629
[10] Li H,Ke H,Ren G et al. Biophys. J.,2009,97:2811
[11] Zhang Y,Li B,Xu Y et al. Plant Cell,2013,25:2504
[12] Li S,Wang R,Li D et al. Sci. Reports,2014,4:4834
本文选自《物理》2020年第10期,经授权转载自微信公众号“中国物理学会期刊网”。
话题:

0
推荐
财新博客版权声明:财新博客所发布文章及图片之版权属博主本人及/或相关权利人所有,未经博主及/或相关权利人单独授权,任何网站、平面媒体不得予以转载。财新网对相关媒体的网站信息内容转载授权并不包括财新博客的文章及图片。博客文章均为作者个人观点,不代表财新网的立场和观点。
 京公网安备 11010502034662号
京公网安备 11010502034662号 {kind=link}