阅读:0
听报道
跑调的人不光唱得跑调,还不知道自己跑调,因为他们压根听不出歌的调调。
撰文 | 哔普星人
人类总能变着法儿地寻找快乐,但万变不离其宗,现代人迷恋的“唱跳rap篮球”根本没出老祖宗们“声色犬马”的传统艺能之窠臼,光怪陆离的活动样式终究只是几种能激活我们伏隔核的快乐模式的缤纷呈现。其中最打头的享乐模式——唱,或者说音乐——毫无疑问占据了人类娱乐生活的顶级流量。
祖宗们留下了不少听起来不那么正能量的音乐相关成语:淫词艳曲、靡靡之音、夜夜笙歌、歌舞升平……除了规劝统治阶级“要励精图治,别沉迷享乐”,也从某种程度上支持了这样一种观点:音乐是一种奇技淫巧的发明,是纯粹的社会创造,就好像哈根达斯只是人类口舌的情人,而非生存刚需一样,不具备进化学层面的意义。
不过很多科学家都不这么认为。他们更愿意从生物学的视角阐释音乐起源。2005年,Science将语言和音乐的进化根源列入未来最具挑战的125个科学问题;到2008年,Nature更是以“科学和音乐”为主旨发表了数篇随笔,从多个学科角度,对音乐是什么、人类为何创作音乐、人类如何创作音乐、人类缘何欣赏音乐、以及音乐的发展变迁等问题进行了概述及讨论。[1-7]
追溯音乐的源头
虽然时至今日,我们还未破解音乐与进化的关系,但越来越多的遗传学和神经科学证据都在逐渐汇集成一份进化音乐谱。
在科学定义中,人类的音乐能力包括两方面:①对节奏、音高、旋律等音乐元素的感知、认知与生产以及②对音乐的生理、心理和情绪反应。
如果音乐能力具有进化意义,那么必然存在一个自然选择机制。例如:完全找不到律动的,淘汰;没有一个音唱在调上的,淘汰;听到一段悲伤G大调后,内心毫无波澜甚至有点想笑的,淘汰。
达尔文于19世纪提出了音乐的性选择假说。他认为人类的音乐能力恰如鸣禽或某些鲸类动物的鸣唱技艺,是影响异性好感度的关键指标之一;乐感基因存在缺陷的走调天王很可能找不到伴侣一起生孩子,断了血脉。[8]

科学家在德国西南部发现了3.5万年前的古人类使用的长笛。[9]
近几十年的研究者既有支持,又有反对该理论的。反对者的理由如下[10-13]:
1. 在战争、宗教和政治这类跟谈恋爱没半毛钱关系的社会活动中,音乐也频频亮相;它们的功能绝对不是求偶。
2. 所有年龄段的异性恋者都能被同性别的音乐家或音乐团体所吸引。
3. 动物性吸引特征往往只存在于两性当中的某一方,且仅在性成熟阶段表现;但人类的音乐能力不因性别有高低,无论老幼皆显露(婴儿期即具备发现和感知旋律与节奏变化的能力)。
4. 上世纪80年代的调查显示,“嗓音”在男人与女人的15项择偶指标里分别排行倒数第二和第一。
5. 前几年一项调查了一万多名双胞胎志愿者的研究显示:音乐能力与交配成功率不存在明显的相关性(交配成功率以性伴侣数量、首次性行为年龄、关系外性行为以及后代数量作为评价指标)。
也有学者提出了——
母婴联系假说:音乐起源于母婴之间的互动,具备帮助他们建立情感联系的功能;
社会凝聚力假说:音乐如同团体内的粘合剂,有助于成员们发扬利他主义,提升集体行动(例如防御猛兽或敌对部落)的效率;
以及语音学习假说、运动声音假说、节奏适应假说等。[14-18]
不过上述理论均存在很多问题,例如母婴联系说和社会凝聚力说都无法解释音乐跨物种存在的现象——要知道很多具备音乐能力的非灵长类动物是不会进行音乐性的母婴互动,也不需要什么良好社会关系的。
另一方面,人类以外的很多动物都显示出了类似的音乐机能。例如:金丝雀、驼背鲸和白臂猿等动物都会“歌唱”;鲸鱼和鸟类的“歌声”与人类的声乐结构相似;黑猩猩喜欢听非洲和印度音乐……这些现象都支持了音乐源于生物进化的说法。
寻找感知音高的脑区
绘制出负责音乐信号处理的脑区图是神经学家们的艰巨任务。
如果我们能非常体系化地阐明大脑的哪些回路用于旋律的感知,又是哪些承担节奏处理的工作,还有哪些决定着我们对音乐情绪的体验,以及这些神经运作具体怎样开展,会受何种因素影响,彼此之间是否存在重叠或协同的情况……那就很可能解答一系列现阶段充满争议的进化起源问题,也将为精准定位音乐基因奠定基础。
研究者们已经大致划出了一些关键区域:颞上沟负责旋律处理,颞上回负责节奏处理,左侧额下回负责音高记忆、音乐句法和情绪;另外,被大脑喜欢的音乐可以刺激伏隔核,触发奖赏系统,带来快感——这也是支持“音乐起源于进化”这一说法的最有力证据。[19-23]
很多学者之所以否定社会凝聚力假说,就在于大脑奖赏回路往往是由最迎合生物本能的刺激(例如性、酒精、赌博等)来激活的,岂是“团队凝聚力”可企及的?
不过以上这些粗略的划分还难以阐释清楚音乐脑机制的特异性。一些科学家试图从音高层面进行更深层的探究。
这里需要介绍一下音高(pitch)的概念。音高分为绝对音高(absolute pitch,简称AP)和相对音高(relative pitch,简称RP)。如果我们说一个人具备绝对音高感,意思就是他能精准判断某声孤零零的叹息处于哪个音阶;只拥有相对音高感的人则必须在参考系里凭借比较和记忆来辨别各个音的高度。
绝对音高感是一种罕见才能,仅0.01%左右的人天赋异禀;史载,拥有绝对音高感的音乐家包括莫扎特、贝多芬、圣·桑等人。一般来说,普通人可以通过恰当训练培养出一定的相对音高感;而那些天赋出色,又动辄玩音乐几十年的专业大佬,往往能有绝对音高的判断力,例如杰伦力宏俊杰等。
与之相对的,是所谓的乐盲(amusia,又称失歌症,约占全球人口的1.5%)——音乐能力存在明显缺陷的群体。乐盲可分为获得性乐盲(acquired amusia)和先天性乐盲(congenital amusia),前者由后天的脑损伤造成,后者则是遗传缺陷所致。
如果按“盲区”的类型划分,可分成音高处理障碍、节奏处理障碍、音乐情绪识别障碍等几种类型。其中最主要的,也是最常被我们提起的,就是音高处理障碍型乐盲。他们难以分辨出旋律里的细微音高差异——用更含蓄点的话来说,他们不光唱歌走调,还不知道自己走调,因为他们压根听不出歌的调调。

“跑调天后”弗洛伦斯·福斯特·詹金斯(Florence Foster Jenkins)是一位热爱演唱的乐盲,凭借超凡脱俗的音乐天赋在上世纪的美国音乐界名噪一时。图左为詹金斯本人,图右为梅丽尔·斯特里普在2016年上映的电影《跑调天后》中饰演的詹金斯。
很多科学家都聚焦于极具音高感和缺失音高感的人群,开展脑区结构研究。
有人通过脑成像技术分析了普通人和擅长判断绝对音高的专业音乐人的大脑结构差异,结果发现:后者的左颞平面(left planum temporale)、颞上回(superior temporal gyrus,STG)、左侧额下回(left inferior frontal gyrus)以及右侧缘上回(right supramarginal gyrus)更大更壮;颞叶结构间存在更为紧密的连接。[24]
加拿大蒙特利尔大学的伊莎贝拉·珀利兹(Isabella Peretz)多年来专攻失歌症研究。
她发现乐盲人群相比正常人,额下回(inferior frontal gyrus,IFG)和听觉皮层的颞上回(前文提到的STG)之间的右侧额颞叶联结网络表现出异常,颞下回(inferior temporal gyrus)和听觉皮层之间的联结减少,连接听觉皮层和额叶皮层的右侧弓状纤维束(arcuate fasciculus)也减少了。
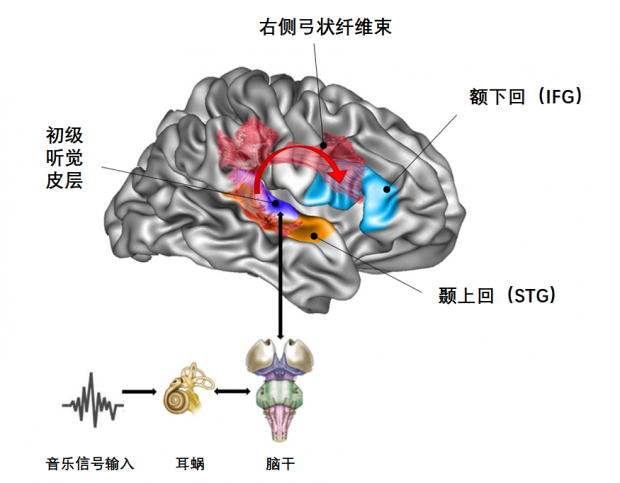
乐盲人群大脑内的特定神经网络存在异常丨图片来源:参考文献[25]
此外,他们还让志愿者聆听同一段旋律,同时借助神经成像技术观察其脑部结构,最后发现听觉皮层并非决定音调感知能力的区域,他们推测是额下回(IFG)与颞上回(STG)之间的正、负反馈的循环运行过程影响着音调信息的传递——但乐盲的大脑难以形成此类反馈,因而无法把握音调。[25-26]
探索音乐才能的基因
证明音乐的生物属性,最根本的方法就是拿出基因证据。
曾有研究者进行过不同人种的音乐能力比较。他们比较了东亚人种(中国人和韩国人)与高加索人种的音高感和节奏感(全部实验对象都是音乐门外汉),发现前者在音高处理方面得分78.2,高于后者的58.2;二者在节奏处理能力上无差距。
这里需要重点说明另一个关键发现:韩国人的音乐表现和中国人水平相当。很多人认为中国人的音高感比金发碧眼的老外强,原因在于汉语富于语调变化,例如普通话有四声,粤语有六个调,闽南语的音调变化甚至多达八个,而英语基本没有明显的语调变化;这使我们对音调更为敏感。但要知道韩语也不存在音调变化,除了遗传因素,我们找不出其他理由解释那些韩国人和我们一样出色的音调判断力。[27]
也有科学家对芬兰人的音乐才能展开过调研。通过测试和分析,他们得出结论:对于这些芬兰人,音高处理能力与节奏处理能力的遗传率(heritability,又称遗传力)分别为57%和21%。所谓的遗传率指的是某一性状受到遗传控制的程度。他们还确定了与音乐能力差异相关的至少46个基因组区域。[28]
另外,珀利兹等人曾分析乐盲的家族聚集情况,发现失歌症人士的一级血亲(父母、子女、兄弟姐妹)也同为乐盲的概率高达39%;这表明该缺陷具有可遗传性。[29]
注:家族聚集是指某一病症、性状或行为在先证者(第一个被发现有问题的家族成员)家族中出现频率高于随机水平的现象。
以上发现只是音乐遗传学视野下的一隅,但也足以令我们隐约窥见那些驱动着人类乐感的基因片段。
音乐的DNA,具体位置在哪儿呢?
我们现在只能给出很多“相关”和“大概”,做不到精准定位。可以先看看下面这张基因清单[30-35]:
ADCY8基因,影响学习和记忆,位于8号染色体长臂24区,被认为与绝对音高相关。
EPHA7基因,关乎神经连接及发育,位于6号染色体长臂16区,被认为与绝对音高相关。
UNC5C基因,负责神经生长因子受体UNC5C表达,位于4号染色体长臂22区,被认为与音高和节奏相关。
TRPA1基因,与内耳毛细胞转导通路相关,位于4号染色体长臂13区到21区,被认为与音高和节奏相关。
AVPR1A基因,影响社会认知和行为,以及空间记忆,位于12号染色体长臂14区到15区,被认为与音高、节奏和音乐记忆相关。
PCDH7基因,负责原钙粘蛋白7的表达,位于4号染色体短臂14区,被认为与音高识别能力高度相关。
ZDHHC11基因,影响丘脑中的5-羟色胺转运体结合潜能,位于5号染色体短臂15区,被认为与音乐创造力相关。
……
不难看出,这些基因似乎都通过影响与音乐能力存在关联的其他因素,例如认知能力或记忆能力,间接左右人体的乐感。我们缺少能给出实锤的决定性基因。
近几年有研究者发现,乐盲可能与Williams-beuren综合征存在关联;该遗传病由于7号染色体上的基因缺失引起。这一发现打开了一条或许能直接引向音乐基因的通途。因为探究决定音乐才能的基因的最有效方法,就是比较缺陷人群与正常人的遗传差异。目前人们就乐盲所掌握的遗传学证据仍然非常有限。

Williams-beuren综合征在人群中的发病率为万分之一。患有此类疾病的人通常会有异于常人的面部结构,可能存在多种健康问题,显示出过度友好的待人态度丨图片来源:
珀利兹说道:“这个方向任务多,难度大,研究方法比神经成像那些东西贵得多;但它有望为理解音乐的生物学基础提供一个全新视角。一切努力和等待都是值得的。” [36]
参考文献
[1] Kennedy D & Norman C. What do we know? Science. 2005; 309: 75.
[2] Ball P. Science & music: Facing the music. Nature.2008; 27: 1180-5.
[3] Barron M. Science & music: Raising the roof. Nature. 2008; 453: 859-60.
[4] Cook N. Science & music: Beyond the notes. Nature. 2008; 453: 1186-7.
[5] Huron D. Science & music: Lost in music. Nature. 2008; 453: 456-7.
[6] McDermott J. Science & music: The evolution of music. Nature. 2008; 453: 287-8.
[7] Zanette D. Science & music: Playing by numbers. Nature. 2008; 453: 988-9.
[8] Darwin C. The descent of man, and selection in relation to sex. 1871.
[9] Conard NJ et al. New flutes document the earliest musical tradition in southwestern Germany. Nature. 2009; 460: 737-40.
[10] Hagen EH & Bryant GA. Music and dance as a coalition signaling system. Human Nature. 2003; 14: 21-51.
[11] Zentner MR & Kagan J. Perception of music by infants. Nature. 1996; 383: 29.
[12] Daniel HJ et al. Values in mate selection: A 1984 campus survey. College Student Journal. 1985; 19: 44-50.
[13] Mosing MA et al. Did sexual selection shape human music? Testing predictions from the sexual selection hypothesis of music evolution using a large genetically informative sample of over 10,000 twins. Evolution and Human Behavior. 2015; 36: 359-66.
[14] Falk D. Prelinguistic evolution in early hominins: Whence motherese? Behavioral and Brain Sciences. 2004; 27: 491-503.
[15] Roederer JG. The search for a survival value of music. Music Perception. 1984; 1: 350-6.
[16] Patel AD. Musical rhythm, linguistic rhythm, and human evolution. Music Perception. 2006; 24: 99-104.
[17] Larsson M. Self-generated sounds of locomotion and ventilation and the evolution of human rhythmic abilities. Animal Cognition. 2014; 17: 1-14.
[18] Wang TY. A hypothesis on the biological origins and social evolution of music and dance. Frontiers in Neuroscience. 2015; 9: 30.
[19] Lee YS et al. Investigation of melodic contour processing in the brain using multivariate pattern-based fMRI. Neuroimage. 2011; 57: 293-300.
[20] Grahn JA & Brett M. Rhythm and beat perception in motor areas of the brain. J Cogn Neurosci. 2007; 19: 893–906.
[21] Watanabe T et al. Memory of music: Roles of right hippocampus and left inferior frontal gyrus. Neuroimage. 2008; 39: 483-91.
[22] Sammler D et al. Are left fronto-temporal brain areas a prerequisite for normal music-syntactic processing? Cortex.
2011; 47: 659-73.
[23] Koelsch S et al. Investigating emotion with music: An fMRI study. Hum Brain Mapp. 2006; 27: 239-50.
[24] Dohn A et al. Gray- and white-matter anatomy of absolute pitch possessors. Cereb Cortex. 2015; 25:1379-88.
[25] Peretz I. Neurobiology of Congenital Amusia. Trends Cogn Sci. 2016; 20:857-67.
[26] Albouy P et al. Impaired pitch perception and memory in congenital amusia: the deficit starts in the auditory cortex. Brain. 2013; 136: 1639-61.
[27] Hove MJ et al. Ethnicity effects in relative pitch. Psychon Bull Rev. 2010; 17: 310-6.
[28] Kanduri C et al. The genome-wide landscape of copy number variations in the MUSGEN study provides evidence for a founder effect in the isolated Finnish population. Eur J Hum Genet. 2013; 21: 1411-6.
[29] Peretz I et al. The genetics of congenital amusia (tone deafness): a family-aggregation study. Am J Hum Genet. 2007; 81: 582-8.
[30] Theusch E et al. Genome-wide study of families with absolute pitch reveals linkage to 8q24.21 and locus heterogeneity. Am J Hum Genet. 2009; 85: 112-9.
[31] Gregersen PK et al. Absolute pitch exhibits phenotypic and genetic overlap with synesthesia. Hum Mol Genet. 2013; 22: 2097-104.
[32] Pulli K et al. Genome-wide linkage scan for loci of musical aptitude in Finnish families: evidence for a major locus at 4q22. J Med Genet. 2008; 45: 451-6.
[33] Ukkola LT et al. Musical aptitude is associated with AVPR1A-haplotypes. PLoS One. 2009; 4: e5534.
[34] Ukkola-Vuoti L et al. Genome-wide copy number variation analysis in extended families and unrelated individuals characterized for musical aptitude and creativity in music. PLoS One. 2013; 8: e56356.
[35] Oikkonen J et al. A genome-wide linkage and association study of musical aptitude identifies loci containing genes related to inner ear development and neurocognitive functions. Mol Psychiatry. 2015; 20: 275-82.
[36] Lense MD et al. (A)musicality in Williams syndrome: examining relationships among auditory perception, musical skill, and emotional responsiveness to music. Front Psychol. 2013; 4: 525.
话题:

0
推荐
财新博客版权声明:财新博客所发布文章及图片之版权属博主本人及/或相关权利人所有,未经博主及/或相关权利人单独授权,任何网站、平面媒体不得予以转载。财新网对相关媒体的网站信息内容转载授权并不包括财新博客的文章及图片。博客文章均为作者个人观点,不代表财新网的立场和观点。
 京公网安备 11010502034662号
京公网安备 11010502034662号 {kind=link}