阅读:0
听报道
进化与癌症如影随形,进化塑造了生命也同样创造了癌症。在生命之树上,从海绵到仙人掌,从狗到大象,科学家都发现了它们身上存在进化的遗迹——癌症。但同时我们也发现了抑制肿瘤的新机制。

本文经授权节选自《狡猾的细胞:癌症的进化故事与治愈之道》(中信出版社)第四章《癌症长满生命之树》,标题为编辑所加,内容有删改。
撰文丨雅典娜·阿克蒂皮斯(亚利桑那州立大学副教授)
翻译丨李兆栋
当乔舒亚·席夫曼(Joshua Schiffman)的伯尔尼兹山地犬被诊断出患有癌症时,席夫曼难以相信这是真的。席夫曼是一名研究癌症的科学家,也是一名儿科肿瘤医生,他自己也是癌症幸存者。让他万万没有想到的是,他自己心爱的宠物也成了他所研究的疾病的受害者,而他自己十几岁的时候就曾患过这种疾病。席夫曼意识到,癌症不仅仅是一种影响人类的疾病,它也影响着生命之树上的许多其他生物。
自己的宠物身患癌症,这样的经历促使席夫曼去更多地了解狗对癌症的易感性,他惊讶地发现它与人类的癌症易感性有许多相似之处。和人类一样,BRCA1/2基因的突变也会增加狗患上乳腺癌和卵巢癌的风险。狗的癌细胞也有TP53基因突变。在人类当中,TP53基因突变会导致一种被称为李–佛美尼的综合征。这是一种遗传性疾病,患者一生中更容易患上癌症。患有慢性髓细胞性白血病的狗甚至被发现携带有BCR/ABL染色体易位,它和我在第三章中讨论的染色体易位是同一个,在人类慢性髓细胞性白血病中非常典型。[1]
狗的癌症和人类癌症的相似性远不只在于遗传风险因素,如BRCA、TP53和BCR/ABL基因的变化。狗和人类患癌症的风险都与较大的体型有关。[2]在上一章中,我们看到因为细胞增殖和控制细胞行为之间的权衡,身体迅速地长大会导致更高的癌症风险。当生物个体行走在发育的平衡钢索上时,它们的细胞必须在过度增殖和过度控制之间取得平衡,个体才能正常发育,成年之后才能成为演化上的适者。因此,如果所有其他因素都一样的话,我们可能会看到体型较大的生物更容易患癌症。
然而其他因素并非都一样——至少大象就是个反例。大象的细胞数量比我们多100倍,然而,它们的癌症发病率却比我们低得多。[3]事实上,如果我们比较不同的物种,我们会发现更大的体型并不会带来更高的癌症风险,这种更高癌症风险与更大体型之间的相关性似乎只在物种之内才会出现。在上一章中我们知道,体型变大会增加癌症风险,是因为需要更多的细胞分裂才能长成一个更大的身体并维持它。那么,为什么在各个物种之间,体型与癌症的风险不存在相关性呢?
在本章中,我将给大家讲述演化是如何解决这个被称为佩托悖论[4]的难题的。我还将仔细探讨一下生命之树上的不同物种对癌症的易感性有所不同的一些原因,从最简单的多细胞生物到大型、复杂的生物,如大象。了解癌症如何影响其他生命形式,以及生命如何演化出抑制癌症的能力,可以帮助我们深入了解我们作为人类为什么容易患上癌症,并对我们制定新的癌症治疗和预防策略提供指导。
癌症遍布不同生命形式
在这本书的开头,我谈到了仙人掌,以及它们由于正常的细胞增殖失去控制而产生的让人叹为观止的各种冠状、多节状以及和大脑一样的生长结构。这些仙人掌令人好奇,因为本质上,它们就是癌症的仙人掌版本——细胞已经成功逃脱多细胞行为的正常限制,开始失控生长。
凤头仙人掌就是缀化植物的一个例子。像仙人掌这样的植物实际上是形态多样的,其生长样式往往有很多不同的面(faces),因此学者用术语“缀化”(fasciation)来描述之。当植物生长尖端的细胞(被称为分生组织细胞)从单个尖端扩张成一排细胞时,就可能出现缀化。这些细胞分裂时,会形成一个扩大的增殖细胞带,这些细胞会长成扇形,有时甚至会折叠起来,形成像人的大脑那样的生长模式。这种缀化现象并不仅仅局限于仙人掌,它在许多其他植物中也会出现。有些花可以发生缀化,长成形状怪异、被拉长的花朵。烟草植物也经常发生缀化,改变其叶子形状和开花的图案。即使是松树这样的大树,也可能会发生缀化,导致树干变得宽阔而沉重,看起来不太稳定。在它们长高的过程中,这些树干会膨胀成扇子的形状。

长了冠的粗枝木麻黄,它没有长出典型的分支结构,而是形成了一个由分化失调的组织组成的大的扇形。丨图源:Forest and Kim Starr/Flickr
多年来,我对仙人掌的兴趣让我迷上了缀化现象,后来进一步扩展到整个生命之树,最终覆盖多细胞生命的所有分支。从技术层面来讲,绿藻也属于植物——包括高大的松树到池塘里的浮渣在内的所有生物体。我和同事们发现,癌症和类似癌症的生命现象在绿藻中会发生,在生命之树的所有其他分支之中也会发生。我们在蛤、昆虫、各种动物、珊瑚、真菌,当然还有植物中都找到了它们存在的证据。
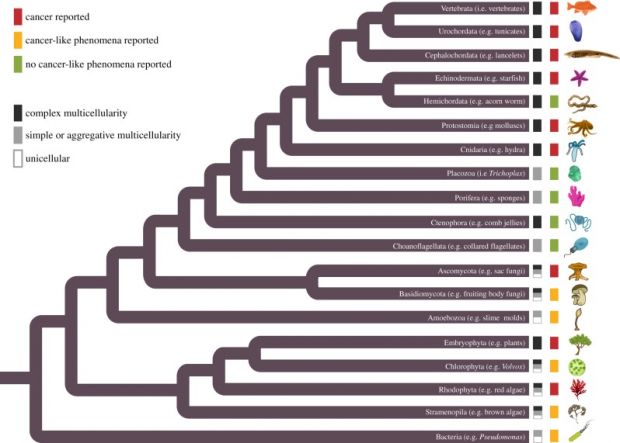
我们发现在多细胞生命的每个分支上都有癌症和癌症样现象(分化失调和过度增殖)的报道。丨图源:参考文献[5]
我们还发现,在所有这些不同物种当中,癌症的发生总是与违背多细胞合作的基本原则有关:对细胞增殖调控的破坏、异常细胞存活、混乱的细胞分工(换句话说,细胞分化过程失控)、营养资源的垄断以及对细胞外环境的破坏。我在前面已经提到,不同物种身上的癌症和类似癌症的现象都可以用细胞作弊这一共同的框架来讨论。与许多其他我们可以使用的定义不同,把癌症定义为细胞作弊可以让我们跨越生物学上各不相同的物种来讨论癌症。
通常来讲,癌症是一个以动物为中心的定义,使用癌细胞的侵袭和转移作为标准。细胞侵袭要求细胞突破其基膜,但并非所有生物的组织都由基膜包裹起来,也并非所有生物都拥有可方便癌细胞转移的循环系统。要定义癌症,将重点放在细胞作弊上会是一个更加通用的方法,这使我们能够在不同的物种之间使用一套广泛适用、与多细胞合作各个部分密切相关的特征。
许多生物学家认为,侵袭性癌症不可能发生在植物身上,因为植物拥有细胞壁和更加固定的组织结构。然而,我们已经看到,植物很容易受到类癌生长(缀化)的影响。这些生长虽然不是侵袭性的,但是它们具有与癌症有关的细胞作弊的所有特征:细胞过度增殖、应当死亡的细胞没有死去、营养资源垄断、细胞分工系统崩溃(花的形状遭到破坏),以及对细胞与细胞之间共享环境的破坏(例如,增加组织死亡的可能性,使整棵植株更容易被感染)。而且,侵袭性生长有时也会在植物身上发生。我们在回顾生命之树上的癌症时,惊喜地发现有一篇论文报道了植物身上的一种侵袭性生长——研究者发现了一排突破现有组织的侵袭性细胞[6]。
这种侵袭性的生长甚至也能够满足对癌症的更严格的传统定义,这表明根据所有检测方法和定义标准来看,植物也可能会得癌症。
目前为止,我们还没有在数据库(由加州大学圣巴巴拉分校的艾米埃米·博迪(Amy Boddy)创建的全面记录癌症的数据库,约17万份动物记录,来自约13000个物种)中发现任何完全不得癌症的动物——每个至少有50份动物记录的物种都至少存在一个肿瘤案例。在撰写本书时,在数据库中发现的癌症发病率最高的物种是白鼬、刺猬和豚鼠。其他癌症发病率非常高的动物包括猎豹和袋獾(甚至在除去袋獾可传染的面部肿瘤的病例的情况下,其发病率依然很高)。
除了建立跨物种癌症风险数据库之外,我们还在研究扁盘动物和海绵——我们发现,纵观所有物种的患癌情况,这些“简单”的生命形式似乎能够抵抗癌症(扁盘动物和海绵不在我们上面讨论的比较肿瘤学数据库中,因为它们通常不是由兽医治疗的,也不是在动物园里治疗的,而数据库里的数据是从这两处获得的)。
我的同事和合作者安杰洛·福尔图纳托(Angelo Fortunato)负责该项目,重点关注这些古老的多细胞生命形式,研究它们抵抗癌症的能力。
福尔图纳托重点研究了几个看似不会得癌症的物种(我们最初的文献综述中没有发现有关这些物种患癌症的报告)。他在实验室中研究的第一批物种包括一种海绵,拉丁学名叫Tethya wilhema,这种生物几乎只是一个由基本上没有分化的细胞组成的集合,身体上有小孔和通道,水和营养物质可以流经它们。福尔图纳托发现,海绵可以说具有极强的抗癌能力;它们可以耐受极高的辐射(会诱导DNA损伤)而没有任何明显的类癌生长。他在观察这些海绵对辐射的反应时注意到,有时它们会收缩几天,然后反弹到以前的尺寸,且没有他能够观察到的明显的奇怪的生长或颜色变化来表明癌症的发生。福尔图纳托目前在运用分子生物学技术来试图揭示导致这种看似能抵抗DNA损伤的适应能力的机制。
他还正在研究丝盘虫(Trichoplax adhaerens,一种扁盘动物)的抗癌能力,这种生物体从理论上来讲属于动物,但其实它基本上就是一袋细胞,外面有一层由细胞组成的外皮来帮助它移动。当福尔图纳托将丝盘虫置于辐射下时,他有时会观察到有些黑暗的区域(可能是癌细胞)在它们体内生长。有时,这些变暗的区域会移动到生物体的外缘,然后似乎被挤压,或被掐掉,黑暗细胞便不复存在。
这可能是一种癌症抑制机制,像扁盘动物这样没有复杂组织和器官系统的生物体利用这种机制来剔除潜在的问题细胞。这种策略似乎只在简单生物身上才会奏效,但如果继续深究,将细胞挤出来并掐掉,对于组织的层面甚至是人类这样的大型生物来讲,看起来也可能是一个可行的策略。例如,在人的结肠中,增殖过多的细胞会被它们的细胞邻居给挤出来。邻近区域的细胞可以产生一系列肌动球蛋白(肌肉的组成成分),组成环状,硬生生地把这些有问题的细胞给挤出来。[7]
福尔图纳托的研究工作让我们稍微了解了简单生物体演化出的保护自己免受细胞作弊所害的方式,同时它也鼓励我们所有人去提出问题,扩大我们的参照系——审视整个生命之树,更好地了解我们的癌症抑制机制如何演化而来。
细胞更多,癌症更多?
在上一节中,我探讨了小而简单的生命形式当中的癌症抑制机制,但是在更大、更复杂的生物(如人类和大象)当中,情形又会如何呢?大型而又复杂的生物如何在足够长的时间里控制住癌症,最终得以成功繁殖呢?
要成为一个多细胞生物,细胞增殖是必不可少的,但它同时也会增加我们对癌症的易感性,因为突变随时都会在分裂增殖的细胞中出现。生物体越大,要长成该大小的体型所需的细胞分裂次数就越多,维持该体型所需的细胞分裂次数也就越多(因为组织需要不断更新)。此外,生物体型越大,在任何时候可能发生变异的细胞的数量也越多。事实上,如果我们看一下某个物种内的癌症发病率,就会发现较大的个体患癌症的风险更高。例如,大型犬类(体重超过约20千克)比体型较小的犬类患癌症的风险更高[8],同样,人类中的高个子患癌症的风险也高于矮个子,每增加10厘米的身高,患癌症的风险就会增加约10%[9]。然而,如果横向比较不同的物种,这种体型越大癌症风险越大的模式就不再成立了[10]。
在这一章开头,我们讲到大象的细胞数量是人的100倍之多,然而它们患上癌症的风险却并不是我们的100倍。以大象这样的体型和寿命,它们对癌症的抵抗力令人吃惊。事实上,与许多体型更小的动物包括人类相比,大象的癌症发病率要低得多。而另一方面,老鼠患癌症的概率要比我们高,尽管它们体型要比我们小得多。这种悖论在寿命方面也存在:寿命越长,细胞分裂并暴露在潜在的诱导突变的因素中的时间就更久,相应的癌症发生的机会就更多。然而,我们比较不同物种之间的癌症风险,就会发现各个物种的癌症发病率和寿命之间并没有什么相关性。
癌症风险与体型和寿命无关,这一现象被称为佩托悖论,由牛津大学的统计流行病学家理查德·佩托爵士(Sir Richard Peto)在20世纪70年代提出。他指出,从细胞的层面比较,人类细胞的抗癌能力一定比小鼠细胞强,否则我们在很小的时候就会死于癌症[11]。我和同事在过去几年中的研究证实了这一现象:寿命更长、体型更大的物种的癌症发病率并不高于寿命较短、体型较小的物种[12]。
生命史的决策
在生命的旅程当中,我们所有人都行走在钢索上,在细胞自由和控制之间取得平衡。给予细胞过多的自由,就会增加我们患癌症的风险,而对细胞管控太多,又会让我们面临生长停滞和演化失败的风险。对于其他多细胞生物来说,同样如此。
大象玩的是长线游戏:它们生育更晚,又没有任何天敌,所以它们在癌症抑制上投入更多,以令自己能够活足够久以获得该策略的回报。这就意味着它们必须要走过一条更长的钢索,才能抵达获得繁殖回报的点。它们不仅要稍微靠右倾斜一点儿以便更有可能最终获得这种繁殖回报,而且如果它们要活到成功繁殖的年纪,还必须整体更好地达到平衡。
使生物向更大的细胞自由度、更混乱、癌症风险更高的一边倾斜的一部分力量来自生物体内部,另一部分力量来自生物体外部,如太阳辐射或化学致突变物质造成的DNA损伤。随着生物种群世世代代的演化,其他因素也会影响到细胞自由和控制之间的这种平衡,如外因导致的高死亡率(由于外在原因比如被天敌猎杀而死亡的可能性)和强烈的性选择(繁殖成功与否受到吸引异性以及与同性竞争的能力的强烈影响)。这些演化压力实际上会选择出采取“左倾”策略的生物,因为如果你活得不够久(或者如果你必须要放弃大量的生殖机会来换取较低的癌症风险),抑制癌症所带来的好处与你为此所要付出的代价相比,就变得微不足道了。
在演化生物学中,这些权衡取舍被称为生命史的权衡取舍,因为它们会影响生物体在其一生中对各种“目标”(如生长、繁殖和生存)进行投资的策略。生命史理论的基本理念是:生物用以实现各个目标来最终提高其繁殖成功率的资源(比如时间和精力)是有限的。在一件事上投入更多资源,留给另一件事的资源就得减少。
我们已经看到,有效的癌症抑制策略往往是有代价的。抑制癌症的能力过强,会影响其他与适应能力相关的特征。这就是尽管生物与癌症一起演化了上亿年,仍然没能彻底抑制癌症的原因之一。我们也已经看到,过度抑制癌症会对生命产生负面影响。
农业中的人工选择为我们理解与演化适应相关的特征和癌症风险之间的这些权衡取舍提供了一个独特的窗口。我们会选择性地繁殖动物来让它们获得某些特性,如产卵和产奶,这种严格的人工选择有时可以产生一些让人意想不到的结果,帮助我们了解某些特征背后的权衡。专门养殖用来产蛋的母鸡就是一例。除了下蛋更多,它们的卵巢癌发病率也更高,可能是因为经过选择,它们的卵巢及其周围组织对细胞增殖更加宽容[13]。
鹿角的季节性快速生长也显示了癌症抑制是一种微妙的平衡。冬天的时候,鹿把鹿角磨掉,然后,鹿角在春季和夏季迅速生长,为秋天的繁殖季节做准备[14]。这些雄鹿的鹿角也更容易生长一种奇怪的类似癌症的东西,被称为鹿角瘤。这些鹿角要极其快速地生长,不仅需要细胞快速增殖,也需要严格控制,以免鹿角的生长完全失控。许多迹象表明,癌症相关信号通路与鹿角快速生长的能力之间存在关联。即使是没有长出鹿角瘤的正常鹿角,其基因表达模式也更像是骨癌(而非正常骨组织)[15]。

此外,肿瘤促进基因在这样的鹿角中也会被表达,基因测序结果显示,与癌症相关的基因(原癌基因)在鹿的祖先中一直受到正向选择[16]。这些鹿角表明,性选择的性状(因为雌性更可能与拥有较大鹿角的雄性交配)可能会增加癌症易感性[17]。
要成为一个体型巨大的生物就需要细胞的增殖能力更强——这是它们长到这么大的体型并维持其体型大小的前提条件,这也意味着它们患癌症的风险更大。但是当我们比较各个不同的物种时,这种关系就不存在了。大象和其他生命史缓慢的生物在演化过程中获得了一些高招,让它们既能够拥有巨大的体型,又可以抵抗癌症。
大象拥有很多额外的肿瘤抑制基因TP53拷贝,使得它们的癌症发病率很低。(而我们人类只有两个TP53拷贝,一个来自母亲,另一个来自父亲。)前文说过,TP53有助于控制细胞增殖,当细胞受损太大而无法修复时,则会诱发程序性细胞死亡。TP53就像基因组的作弊探测器,监测细胞的异常行为,并做出相应的反应。TP53 只是众多肿瘤抑制基因中的一个,但它是最重要的一个:它能通过监测DNA损伤这样的异常情况,来帮助细胞维持健康状态。如果它检测到损伤,就会停止细胞分裂周期,直到问题解决。如果出现的问题无法解决,TP53 将启动细胞自杀程序,开启一系列的信号转导,最终导致细胞凋亡。由于TP53拷贝的额外存在,大象的这些癌症抑制功能也格外强大:它们对DNA损伤特别敏感,因此当损伤发生时,它们的细胞更容易启动自毁程序。研究分析显示大象有40个TP53基因拷贝,大象细胞在辐射照射下很容易自毁[18]。
大象并非唯一为了让体型更大而演化出癌症抑制系统的生物。北亚利桑那大学的演化生物学家马克·托利斯(Marc Tollis,也是我们研究小组的成员)发现座头鲸基因组中存在凋亡基因的重复。与体型较小的鲸目动物(包括抹香鲸、宽吻海豚和虎鲸)相比,座头鲸在演化过程中更偏向于负责细胞周期控制、细胞信号传递和细胞增殖的基因[19]。
调节与受控
细胞自由和细胞控制之间的平衡是一个动态的过程,终生存在。像TP53这样的基因并不会不停地表达蛋白质——如果这样,它会把走钢索的我们推得太靠右,也会产生一定的代价(如过早衰老或生育能力低)。大象不仅平衡更倾向于右侧,与体型小的动物相比,大象在保持平衡方面也更小心、更积极。体型大而又长寿的动物需要更强的癌症抑制系统和更精细的调控机制,以使生物体在其寿命期限之内保持在这条钢索上的平衡。这不仅仅是表达更多的基因产物、提高细胞控制强度的问题,也是一个需要在正确的时间表达适量的基因产物的问题,这样才能保障基因产物表达平衡,否则生物体细胞将陷入混乱。
那么生物体如何调控那些调控因子呢?一个方法就是构造基因网络(基因之间的连接,使它们可以受到彼此状态的影响),包括从促进细胞自由到促进细胞控制之间的所有基因。通过监测和影响基因产物的表达,这些网络可以帮助生物体在更长的时间里保持这种微妙的平衡(正如我们前面看到的TP53的信号监测功能)。
主要负责让平衡偏向细胞自由(促进细胞增殖)的基因是最古老的基因,它们早在单细胞生物时期就出现了。而主要负责让平衡偏向细胞控制的基因是在单细胞向多细胞过渡的时期演化出现的。这些基因中的许多有时被称为看护基因,它们有助于加强细胞之间的合作,使多细胞有机体成为可能。但还有另一类基因:处在促进自由的“单细胞”基因和加强控制的“多细胞”基因之间的基因[20]。这些基因被称为守门基因,有助于保持整个系统的平衡、动态地响应变化,根据需要分别向平衡的两侧发送信号来进行调整。
“单细胞”和“多细胞”基因之间的守门基因从演化时间上来讲是最新的[21],它们令像人类和大象这样的大型、长寿的生命形式得以平衡细胞自由和细胞控制在整个生命周期中的相互冲突,使它们互相妥协。这些基因可以帮助生物体动态地管理不断变化的力量,否则这些力量就会让本来就很困难的走钢索变得更加不稳定。
参考文献
[1] Joshua D. Schiffman and Matthew Breen, “Comparative Oncology: What Dogs and Other Species Can Teach Us about Humans with Cancer,” Philosophical Transactions of the Royal Society of London, Series B: Biological Sciences 370, no. 1673 ( July 2015).
[2] J. M. Fleming, K. E. Creevy, and D. E. L. Promislow, “Mortality in North American Dogs from 1984 to 2004: An Investigation into Age-, Size-, and Breed-Related Causes of Death,” Journal of Veterinary Internal Medicine 25, no. 2 (March 2011): 187–98; Jane Green et al., “Height and Cancer Incidence in the Million Women Study: Prospective Cohort, and Meta-Analysis of Prospective Studies of Height and Total Cancer Risk,” Lancet Oncology 12, no. 8 (August 2011): 785–94; Sara Wirén et al., “Pooled Cohort Study on Height and Risk of Cancer and Cancer Death,” Cancer Causes and Control 25, no. 2 (February 2014): 151–59.
[3] Lisa M. Abegglen et al., “Potential Mechanisms for Cancer Resistance in Elephants and Comparative Cellular Response to DNA Damage in Humans,” Journal of the American Medical Association 314, no. 17 (November 2015): 1850–60.
[4] R. Peto et al., “Cancer and Ageing in Mice and Men,” British Journal of Cancer 32, no. 4 (October 1975): 411–26; Richard Peto, “Epidemiology, Multistage Models, and Short-Term Mutagenicity Tests,” International Journal of Epidemiology 45, no. 3 (1977): 621–37.
[5] C. A. Aktipis et al. “Cancer across the Tree of Life: Cooperation and Cheating in Multicellularity.” Philosophical Transasctions of the Royal Society B: Biological Sciences 370 (2015).
[6] G. N. Agrios, Plant Pathology (Boston: Elsevier Academic Press, 2005), 922; Philip R. White and Armin C. Braun, “A Cancerous Neoplasm of Plants: Autonomous Bacteria-Free Crown-Gall Tissue,” Cancer Research 2, no. 9 (1942): 597–617.
[7] George T. Eisenhoffer et al., “Crowding Induces Live Cell Extrusion to Maintain Homeostatic Cell Numbers in Epithelia,” Nature 484, no. 7395 (April 2012): 546–49.
[8] Fleming, Creevy, and Promislow, “Mortality in North American Dogs from 1984 to 2004.”
[9] Green et al., “Height and Cancer Incidence in the Million Women Study”; Wirén et al., “Pooled Cohort Study on Height and Risk of Cancer and Cancer Death.”
[10] Leonard Nunney et al., “Peto’s Paradox and the Promise of Comparative Oncology,” Philosophical Transactions of the Royal Society of London, Series B: Biological Sciences 370, no. 1673 (July 2015).
[11] Peto, “Epidemiology, Multistage Models, and Short-Term Mutagenicity Tests.”
[12] Abegglen et al., “Potential Mechanisms for Cancer Resistance in Elephants and Comparative Cellular Response to DNA Damage in Humans”; A. F. Caulin and C. C. Maley. “Peto’s Paradox: Evolution’s Prescription for Cancer Prevention.” Trends in Ecology and Evolution 26, no. 4 (February 2011): 175–82.
[13] Boddy et al., “Cancer Susceptibility and Reproductive Trade-Offs”; P. A. Johnson and J. R. Giles, “The Hen as a Model of Ovarian Cancer,” Nature Reviews Cancer 13, no. 6 (2013): 432–36.
[14] Robert A. Pierce II, Jason Sumners, and Emily Flinn, “Antler Development in White-Tailed Deer: Implications for Management,” University of Missouri Extension, January 2012
[15] Yu Wang et al., “Genetic Basis of Ruminant Headgear and Rapid Antler Regeneration,” Science 364, no. 6446 ( June 2019).
[16] Wang et al., “Genetic Basis of Ruminant Headgear and Rapid Antler Regeneration.”
[17] Boddy et al., “Cancer Susceptibility and Reproductive Trade-Offs.”
[18] Abegglen et al., “Potential Mechanisms for Cancer Resistance in Elephants and Comparative Cellular Response to DNA Damage in Humans.”
[19] Marc Tollis et al., “Return to the Sea, Get Huge, Beat Cancer: An Analysis of Cetacean Genomes Including an Assembly for the Humpback Whale (Megaptera Novaeangliae),” Molecular Biology and Evolution 36, no. 8 (August 2019): 1746–63.
[20] Anna S. Trigos et al., “Altered Interactions between Unicellular and Multicellular Genes Drive Hallmarks of Transformation in a Diverse Range of Solid Tumors,” Proceedings of the National Academy of Sciences of the United States of America 114, no. 24 (June 2017): 406–11.
[21] Trigos et al., “Altered Interactions between Unicellular and Multicellular Genes Drive Hallmarks of Transformation in a Diverse Range of Solid Tumors.”
话题:

0
推荐
财新博客版权声明:财新博客所发布文章及图片之版权属博主本人及/或相关权利人所有,未经博主及/或相关权利人单独授权,任何网站、平面媒体不得予以转载。财新网对相关媒体的网站信息内容转载授权并不包括财新博客的文章及图片。博客文章均为作者个人观点,不代表财新网的立场和观点。
 京公网安备 11010502034662号
京公网安备 11010502034662号 {kind=link}