阅读:0
听报道
新型冠状病毒从哪里来?为何如此难以防控?会在感染者之间交叉传播,变异成更厉害的病毒吗?本文从病毒演化、毒力变化的角度给出了回答。
撰文 | 李庆超(山东师范大学)
2019年底暴发的新型冠状病毒肺炎(Novel coronavirus pneumonia,NCP)是中国自2003年SARS以来最严重的新发病毒传染病疫情,它不仅严重威胁着国民的生命安全,也对我国的经济、社会发展造成了极大的影响,引起了社会各界的高度重视。我们不禁要问,新型冠状病毒(Novel Coronavirus,2019-nCoV,国际病毒分类委员会(ICTV)将新型冠状病毒命名为SARS-CoV-2,目前学界尚有争议,中国专家呼吁命名为HCoV-19,我们暂用大家所熟悉的新型冠状病毒这一名称)是怎么来的,接下来疫情会如何发展,为了避免新的疫情发生,我们应当做些什么?知己知彼百战百胜,回答这些问题,我们就要了解新发病毒从何而来,感染人体后会如何演化发展,病毒的演化给防治病毒带来哪些挑战等一系列问题。接下来,我们从演化的角度认识一下病毒。
在讨论病毒演化之前,我们先做一个澄清,“Evolution”一般翻译为“进化”,其实更准确地说,应该翻译为“演化”。“进化”一词毫无疑问是带有主观褒义色彩的,它暗示了从简单到复杂、从低级到高级等等思考或判断,认为演化一定是沿着某一“光明的未来”前进的。事实上,演化是没有主观意愿、没有价值判断、没有智慧设计的中性过程,它的概念是“连续几代生物种群遗传特性的变化”,这种变化经历自然选择,从而表现为某一特定结果:适者生存。适应性(adaptation)理论上可以用适应度(fitness)来衡量,适应度是指某个群体内某种基因型机体存活并将其基因传递至下一代的相对能力(与其他基因型相比的比较优势) 。适合度越大,存活和生殖机会越高。而什么是“适者”取决于自然选择的具体条件。通常情况下,人们习惯使用“进化”一词也是没有问题的。
病毒的起源及演化研究
病毒演化是病毒研究的重要领域。
由于病毒本身是一类细胞内专性寄生的生物,因此对病毒演化起源的讨论主要集中在病毒与细胞生物起源之间的关系上,主要有三个假说[1](见图1):
1)优先假说:病毒独立、且先于细胞生物起源。
2)退化假说:病毒起源于细胞生物,是某些细胞生物退化而来。
3)逃逸假说:病毒是某些细胞生物的一部分,脱离细胞后形成独立的复制元件。
最近比较新的观点还有协同演化假说和嵌合起源假说,均是以上三种假说的改良或结合[1]。鉴于病毒具有多样不同的基因组种类和颗粒结构,不同的病毒可能具有不同的演化起源,比如RNA病毒可能独立演化而来,而DNA病毒可能起源于细胞生物。

图1. 病毒起源的三种假说[1]。
病毒演化研究主要是通过病毒的基因组序列分析进行的,基因组结构类似或序列具有同源性,则病毒具有亲缘关系,序列越接近,亲缘关系越近,复制周期、宿主范围、致病性、免疫原性则往往越相似。从大的时间尺度来讲,由于病毒与细胞生物之间存在广泛的遗传信息交流,病毒在细胞生物演化中发挥巨大的作用[3],病毒演化研究能帮助我们认识病毒圈“virosphere”这一生命圈中极具多样性的遗传信息库[2],且有利于研究重要的病毒,特别是致病病毒的演化发展。从较短的时间尺度来看,病毒序列差异性及其演变是流行性感冒(流感病毒)、艾滋病(AIDS)和肝炎(例如HBV、HCV)等病毒性疾病流行病学研究的重要方面,同时也是疫苗设计、病毒耐药性研究的理论基础。
病毒变异
病毒演化过程中遗传特性的差异是由病毒变异提供的。
病毒变异的研究,重头戏是致病病毒抗原性变化。“病毒抗原变异“有两个专有概念,分别为抗原漂移(antigenic drift)和抗原漂变(antigenic shift),前者是指较为细微的变化,而后者是指抗原性的巨大变化。
事实上,病毒变异可能发生在病毒基因组的任何位置,其影响的表型也不局限于抗原性。从病毒整个基因组上来说,病毒变异可以分为1)碱基突变(mutation),包括单碱基或小片段的插入或缺失;2)重组(recombination),即同一种病毒不同病毒株间基因组分子的大片段交换;3)重排(reassortment),即分节段基因组病毒的基因分子重新组合。这三种病毒变异的发生机制是完全不同的。碱基突变是病毒基因组复制酶的错误复制导致的,重组是分子间重组交换或模板转移导致的,而重排则是病毒包装过程中分节段基因组选择分配导致的。将病毒基因组比喻为一本书,复制酶相当于抄写员,碱基突变类似于在抄写过程中,抄写员走神,抄错了字符;重组相当于抄写员抄写过程中写到一半换了一本书,一本书上半部分是《西游记》,下半部分是《红楼梦》;而重排则类似于将1~4册《哈利波特》与5~7册《纳尼亚传奇》装在一起。从这个类比中大家不难看出,碱基突变是所有病毒基因组复制中都可以出现的一种变异形式,它的发生频率主要取决于抄写员病毒基因组复制酶的保真性,对于RNA病毒来说,其RNA复制酶不具备校对活性,因此RNA病毒的突变率是最高的,DNA病毒的复制酶具有校对活性,因此突变率较低。而重组和重排则是在特定条件下才能发生的,如同抄写员面对两种以上的书,同一细胞里面感染了两种或以上亲缘关系相近的病毒,才有可能发生重组和重排。
需要指出的是,对于碱基突变来说,由于它是病毒基因组复制酶的错误复制导致的,这种错误是随机发生的,因此病毒的突变可以发生在任何位点上。那么我们在检测病毒序列的时候,是否可以看到突变在整个基因组中随机分布呢?
病毒的突变不能影响病毒复制所必需的蛋白质的编码以及RNA或DNA元件序列,因此,大量的突变往往是有害的,这些含有有害突变的病毒不能复制自身,从而被淘汰了。有害突变之外,剩余的突变还有相当一部分属于“同义突变”,即突变前后对病毒没有影响。最终,只有少数突变对病毒适应性具有正面作用,被富集和固定下来。由于病毒基因组一般较短、信息冗余度很低,囿于病毒自身生存复制需求、宿主先天性免疫和适应性免疫压力的筛选,病毒基因组的突变位点大部分集中在病毒基因组的某些区域,因此我们能够检测到的病毒突变在基因组上的分布是不均匀的。
具体到冠状病毒来说,在病毒刺突膜粒(spike peplomers,S)基因上的某些高变区,可以检测到更多的突变,这些突变往往影响了病毒对宿主受体的亲和力,以及病毒的免疫原性(引发机体免疫保护反应的性能),有可能导致病毒获得感染新宿主的能力、逃逸适应性免疫保护的能力等新的能力。本次暴发的新型冠状病毒的S蛋白存在与其他蝙蝠冠状病毒所不同的四个关键氨基酸点突变,与SARS-CoV具有相似的受体结合能力,导致其获得了感染人的能力[11]。
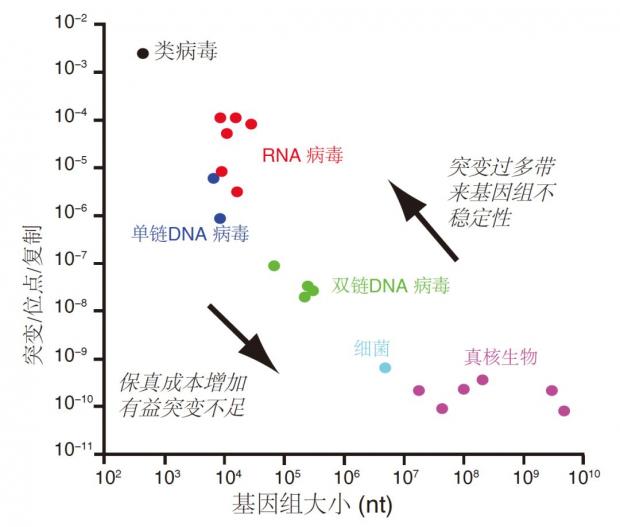
图2. 生物基因组与其变异频率的关系:基因组越短,其变异能力越强,基因组越大,对保真性要求越高。病毒属于快速演化的一类生物,为了降低保真性成本以及积累有益突变,突变率往往很高,其基因组远小于细胞生物[4]。
病毒演化的“加速器”:人类与野生动物遭遇及宿主跃迁
新发病毒一般来自于野生动物,当人与野生动物的生存边界被打破后,两者之间产生本不该发生的接触,给病毒创造了感染人的机会。
动物病毒与原宿主之间都经历了长期的共同演化过程,一般不会造成严重的疾病或死亡,且大多数动物病毒不能感染人,但是少数的动物病毒感染人之后可以造成严重的疾病和高致死率。目前发生过的冠状病毒造成严重感染有:严重急性呼吸综合征(Severe acute respiratory syndrome,SARS)、中东呼吸综合征(Middle East Respiratory Syndrome,MERS)和2019年新型冠状病毒肺炎,包括HIV、埃博拉病毒以及三种冠状病毒在内的新发病毒病都是可以感染人和动物的人畜共患病(zoonosis)。因此野生动物是新发病毒的重要来源。
无论是理论分析还是分子流行病学检测,种种迹象表明,蝙蝠是病毒的存储库[5, 6]。一项研究表明,蝙蝠中的冠状病毒携带率为8.6%,远远高于非人灵长类的0.12%和啮齿类动物的0.32%,且蝙蝠中冠状病毒的种类也多达91种[7]。蝙蝠之所以成为理想的“病毒库”,是因为其具有较长的寿命、较大的活动范围、群居习性以及特殊的免疫系统等特征,使其能够接触广泛的病毒源并且能够长期携带病毒。而冠状病毒可以通过果子狸、单峰骆驼、穿山甲(穿山甲是否为中间宿主,目前存在争议,穿山甲携带的冠状病毒与2019-nCoV差异较大[14])等中间宿主传播给人。目前已知与2019-nCoV亲缘关系最近的是菊头蝠体内发现的RaTG13病毒,与2019-nCoV相似度96%以上,只有近1100个核苷酸的差异,根据病毒之间核苷酸的差异和其他冠状病毒的假定突变率得出的估计,这两种病毒在25到65年前有一个共同的祖先。因此,RaTG13病毒突变为2019-nCoV可能需要几十年的时间,但是由于两者之间存在不同的蛋白酶切位点,推测RaTG13也不是2019-nCoV的直接祖先[15]。另外,目前认为2019-nCoV不是通过重组产生的新病毒[16]。因此,2019-nCoV的直接来源尚需进一步研究。
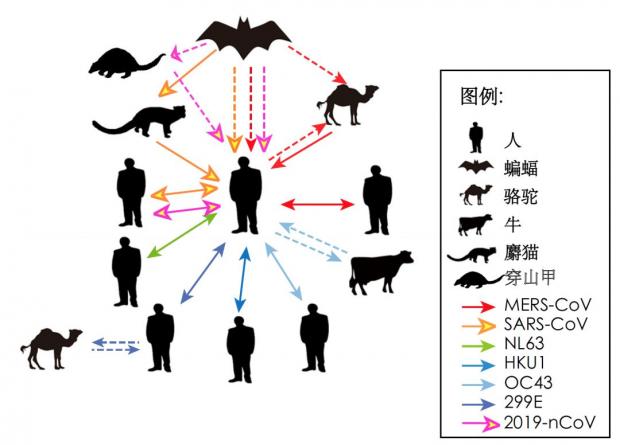
图3. 冠状病毒的种内及种间传播:三次冠状病毒爆发均指向蝙蝠这一自然宿主,及作为野味或者与人密切接触的中间宿主。实线为已证实的传播途径,虚线为疑似传播途径。改编自参考文献[8]。
某些病毒的宿主范围本身就不是特别专一,本身就能够机会性地感染人;而野生动物携带的病毒在不断演化的过程中也可能发生跨物种传播,获得传染人的能力——虽然这种可能性概率极低,但不可忽略。另一方面,冠状病毒的重组突变发生机制虽不明确,但是序列分析发现,冠状病毒之间确实发生过重组。因此说,存在于野生动物中的冠状病毒是有可能通过重组的方式获得新的遗传特性的。我们假设,这些病毒存在于深山老林的野生动物身上,远离人类社会,本是不会引起人类感染的。而环境破坏后,在捕杀、贩卖或宰杀、食用野生动物的过程中,人与野生动物的生存边界被打破,造成的两者之间本不该发生的接触,给病毒发生宿主跃迁、传播人创造了条件。因此,我国出现的两次冠状病毒感染都发生在野味交易市场及其消费链条之上,而禽流感和猪疱疹病毒感染则容易发生在活禽市场、养殖场等高危场所或兽医等高危人群中。
动物宿主在病毒演化中起着关键作用,最著名的例子当属流感病毒的演化:不同的流行性感冒病毒可以感染不同的动物,而有些动物可以被多种流感病毒所感染。
流感病毒含有8个RNA片段的分节段基因组,如果细胞感染了两种不同的流感病毒,则两种病毒的RNA都会在细胞核中复制。当新的病毒颗粒在质膜上组装时,在原有的两套基因组中随机选择8条进行组合而产生新的病毒基因组。从两个亲本遗传RNA产生的新病毒称为重配子,这个过程就是重排。哺乳动物流感和禽流感病毒之间RNA片段的重排交换能够引起流感大流行,例如,2009年H1N1流行毒株是禽、人和猪三种流感病毒的重配体。而在这个过程中,起关键作用的就是猪——鸡、鸭可以感染来自野鸟的禽流感,并传染给猪,猪也可以感染人的流感病毒,猪同时感染两种或两种以上流感病毒,并在体内发生基因重排,就会产生能感染人的强毒力流感病毒。因此,猪成为流感病毒的混合器,加速了病毒演化过程。
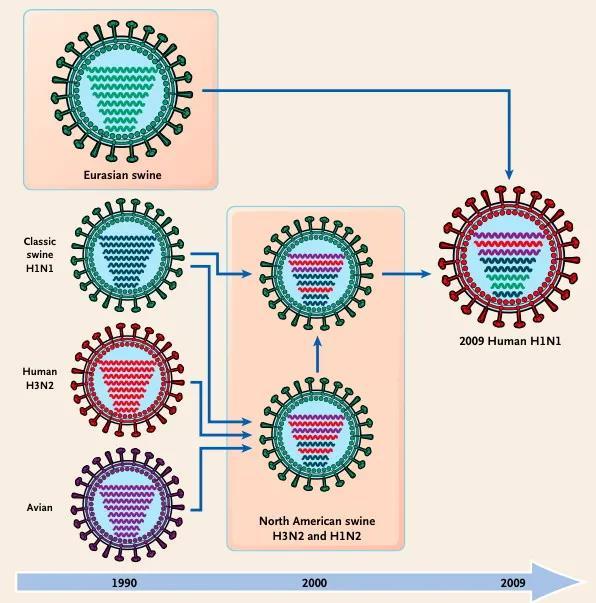
图4. 流感病毒在猪体内重排并造成人类大流行:禽流感、猪流感及人流感病毒均能感染猪,猪在同时感染两种以上的流感病毒后,重排产生新的流感大流行毒株[9]。
病毒的毒力及演化方向
病毒进入感染新宿主后,会优化毒力,增强适应性。
我们可以从两个维度来看病毒的危害性:一,传播率(Transmission rate);二,毒力(virulence)或致死率(Mortality)。在长期演化过程中,毒力和病毒传播能力之间存在权衡关系。毒性强的病毒对其宿主造成很大损害,并产生大量病毒后代,传播能力也很强。但是,如果宿主的疾病限制了宿主的活动性,使其不能与新宿主接触,则该病毒实际上不能有效地感染新宿主,反而具有相对较低的演化适应性。相反,毒性较低的病毒可能感染更多的宿主,因为这些宿主状态足够好,可以与许多其他潜在的宿主接触,但是其毒力必须要能够产生足够多的子代病毒并传播给新宿主。
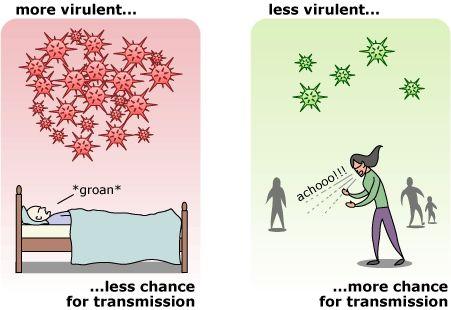
图5. 毒力与传播率的权衡关系:过高的毒力导致宿主无法接触新的潜在宿主,限制了其传播能力;而毒力适中的病毒虽然产生较少的病毒,但因其宿主活动性不受限,可以感染更多新宿主,从而获得更高的适应性[18]。
毒力(virulence)是一个比较笼统的概念,指的是微生物感染宿主和对宿主造成损伤的能力,是由病原体、宿主和环境因素共同决定的复杂特征。单从病毒方面考虑,病毒的毒力可笼统地定义为其致病率或致死率。自然选择发生在病毒的宿主身上,以及病毒传播的过程中,衡量病毒演化的标准是病毒的适应度,而不是毒力。对毒力演化的直接解释是,自然选择会“优化”毒力水平,使病原体适应度最大化。
适应度可以笼统地表示为基本传染数(R0)。在流行病学上,基本传染数R0(Basic reproduction number)是指在没有外力介入,同时所有人都没有免疫力的情况下,一个感染到某种传染病的人,会把疾病传染给其他多少个人的平均数(R0<1,病毒逐渐消亡;R0=1,病毒局部流行;R0>1,病毒流行扩散)。

图6. 基本传染数R0:R0<1,病毒逐渐消亡;R0=1,病毒局部流行;R0>1,病毒流行扩散。来源:电影《传染病》剧照
病毒生存演化的方向应当是通过“优化”毒力“追求”更高的适应性,即R0的最大化,而所谓毒力的“优化”,可能是毒力增强,也可能是毒力减弱。对于某个病毒而言,自然选择到底是增加还是减少病原体的毒力,具体取决于宿主、病毒和环境之间的特定组合。因此,病毒的演化不必然带来病毒毒力的增强,但毋庸置疑的是,毒力增强的病毒演化事件更易引起人们的注意[10]:
1. 口服脊髓灰质炎疫苗(OPV):OPV是脊髓灰质炎病毒的减毒形式,偶尔会恢复为强毒形式并引起脊髓灰质炎暴发。
2. 西尼罗病毒:1999年,西尼罗病毒 NS3蛋白T239P突变产生对鸦科动物的高致死率毒株。
3. 鸡马立克氏病病毒:1960年代第一代鸡马立克氏病疫苗只能对野生病毒感染提供部分的保护效果,减轻疾病症状。疫苗使用大约十年后,出现了毒力更强的野生毒株,人们不得不研发第二代疫苗,而改用新疫苗后,在更短的时间内再次出现了新的强毒力野生毒株。
4. 寨卡病毒:2007年前仅在亚非地区发现轻微感染病例,而2013年南美暴发了具有神经毒性的寨卡病毒,研究发现其由亚洲寨卡病毒演化而来,S139N位点突变可能与其毒力增强有关。
5. 埃博拉病毒:埃博拉病毒A82V突变点可能通过提高对人NPC1受体的结合来增加对人的感染力,且与病例数增加和死亡率增加具有很强的关联。但目前的动物实验研究尚未证实这一点。
6. 兔粘液瘤病毒:强毒力粘液瘤病毒曾被用来抑制欧洲及澳洲的野兔泛滥,该病毒在早期引起大量野兔死亡,幸存的野兔对病毒具有一定的抵抗能力,病毒表现为弱毒力,但是在病毒演化过程中反复出现强毒力的病毒株,平均毒力表现为缓慢上升的趋势。
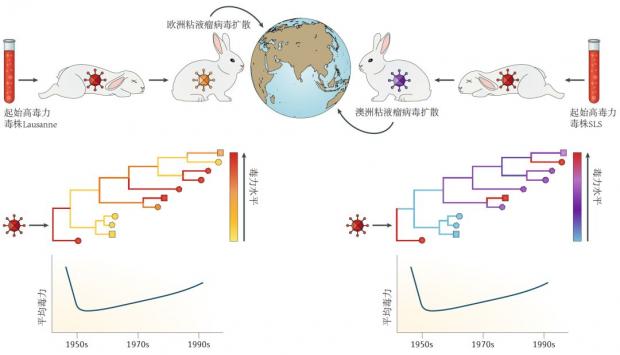
图7. 兔粘液瘤病毒的演化:强毒力粘液瘤病毒被用于治理兔患,病毒短期内毒力下降,但长时间内平均毒力增加[10]。
冠状病毒的变异及演化
冠状病毒具有RNA病毒的中高水平的突变率,且存在重组的可能性,但短期内不是我们所需要关注的重点。
这是因为,冠状病毒的基因组是单股正链RNA,不存在重排的可能性。碱基突变方面,有研究报道,冠状病毒的RNA复制酶具有外切核糖核酸酶活性,对病毒基因组复制和亚基因组合成起到一定的纠错功能[13],但是,研究表明,冠状病毒碱基替换率与其他单股RNA病毒相比,处于中到高水平,约为10-4替换/年/位点[8],也就是说一年时间内,病毒复制过程中在某一位点发生突变的概率是一万分之一。此外,序列分析发现,不同的冠状病毒基因组之间存在重组现象,这种重组既发生在感染人的冠状病毒之间,也存在于动物冠状病毒之间[8]。
最新的研究报道称,2019-nCoV的R0为3.77,高于早期2.2的估计值[12],这说明,与SARS-CoV相比,本次暴发的新型冠状病毒传播能力更强、潜伏期长、携带者症状不典型,主要通过飞沫传播,且最近发现还可能通过口-粪途径传播,给防疫带来很大的困难。病毒的传播方式与其细胞受体在人体的分布、病毒释放途径、病毒在体外的环境抗性和病毒传播所需最低载量有关。最近有报道讨论新型冠状病毒气溶胶传播或者“远程空气传播”的可能性,笔者认为,咳嗽或喷嚏产生的气溶胶体积较小,病毒载量本身就很小,且气溶胶极易干燥,而干燥会极大地降低冠状病毒感染力,因此冠状病毒通过气溶胶进行远程传播的几率极小,因此在旷野活动或者家庭开窗通风是安全的。需要注意的是,从香港“康美楼”案例来看[17],病毒可能通过排粪管进行跨楼层的传播,因此要注意下水道系统的密闭性。
由于医疗资源有限,国家建设了一批方舱医院对感染病患进行救治。不少读者关心,这种环境是否会促进病毒演化,可能导致疫情加重?
首先,我认为大可不必从冠状病毒演化这个角度来质疑方舱医院的安全性,因为病毒的碱基突变变异方式是一种自然发生的过程,如果没有诱变剂的存在,其演化速率是不会发生变化的。另外,短期内被感染的病患可能携带的是相同的病毒株,不大存在新冠病毒之间发生重组产生新毒株的可能性。笔者认为,应当在方舱医院内注意除COVID-19之外的其他传染病的防治工作,避免病人发生多重感染加重病情,并应当注意病人在这种高度密集特殊的环境下产生的心理压力积累。
结 语
COVID-19本身是一种新发病毒病,由于2019-nCoV具有传播能力强、病毒携带者症状隐秘等特点,给防控造成很大困难,如果要让2019-nCoV像SARS-CoV一样销声匿迹,恐怕要付出更大的努力。笔者认为,与流行性感冒病毒相比,新型冠状病毒由于缺乏与人类关系密切的自然病毒宿主,可能不会发展成为一种季节性的流行病。长远来看,如果无法完全清除2019-nCoV,随着人们对2019-nCoV的认识不断加深,治疗经验积累,死亡率大大下降,我们可以将其视作一般的流行感冒,研发药物或疫苗加以防治。如果今后需要研发COVID-19药物或者疫苗,病毒变异是必须考虑的问题,因为病毒变异会导致其药物敏感性的变化,以及抗原性的变化,从而影响药物治疗效果和疫苗保护效果。即便战胜了2019-nCoV,也不排除另一种新的冠状病毒引发另一次新发病毒病暴发的可能性。野生动物是新发病毒病的主要来源,其迁徙不定,无法进行管理;而目前环境破坏、森林砍伐、城市化、长途航空旅行增加、动物(包括牲畜)与人群国际间交流频繁,导致人与野生动物频繁接触和遭遇……这些因素都给传染病防控带来了很大的困扰。野生动物携带各种病原微生物是正常现象,无论是为了保护野生动物,还是为了保护人类健康,都不应该捕杀和食用野生动物。经历了SARS和这次新冠状病毒肺炎疫情后,我们应当吸取教训,坚决抵制捕杀野生动物,绝不食用野生动物。
参考文献
[1] Krupovic, M., Dolja, V.V., and Koonin, E.V. (2019). Origin of viruses: primordial replicators recruiting capsids from hosts. Nat. Rev. Microbiol. 17, 449–458.
[2] Shi, M., Lin, X.-D., Tian, J.-H., Chen, L.-J., Chen, X., Li, C.-X., Qin, X.-C., Li, J., Cao, J.-P., Eden, J.-S., et al. (2016). Redefining the invertebrate RNA virosphere. Nature 540, 539–543.
[3] Emerman, M., and Malik, H.S. (2010). Paleovirology--modern consequences of ancient viruses. PLoS Biol 8, e1000301.
[4] Holmes, E.C. (2011). What does virus evolution tell us about virus origins? Journal of Virology 85, 5247–5251.
[5] Brook, C.E., and Dobson, A.P. (2015). Bats as “special” reservoirs for emerging zoonotic pathogens. Trends Microbiol. 23, 172–180.
[6] Daszak, P. (2017). Host and viral traits predict zoonotic spillover from mammals. Nature 546, 646–650.
[7] Anthony, S.J., Johnson, C.K., Greig, D.J., Kramer, S., Che, X., Wells, H., Hicks, A.L., Joly, D.O., Wolfe, N.D., Daszak, P., et al. (2017). Global patterns in coronavirus diversity. Virus Evol 3, vex012.
Olival, K.J., Hosseini, P.R., Zambrana-Torrelio, C., Ross, N., Bogich, T.L., and
[8] Su, S., Wong, G., Shi, W., Liu, J., Lai, A.C.K., Zhou, J., Liu, W., Bi, Y., and Gao, G.F. (2016). Epidemiology, Genetic Recombination, and Pathogenesis of Coronaviruses. Trends Microbiol. 24, 490–502.
[9] Trifonov, V., Khiabanian, H., and Rabadan, R. (2009). Geographic dependence, surveillance, and origins of the 2009 influenza A (H1N1) virus. N Engl J Med 361, 115–119.
[10] Geoghegan, J.L., and Holmes, E.C. (2018). The phylogenomics of evolving virus virulence. Nat. Rev. Genet. 19, 756–769.
[11] Xu, X., Chen, P., Wang, J., Feng, J., Zhou, H., Li, X., Zhong, W., and Hao, P. (2020). Evolution of the novel coronavirus from the ongoing Wuhan outbreak and modeling of its spike protein for risk of human transmission. Sci. China Life Sci. 382, 1993–1994.
[12]
[13] Minskaia, E., Hertzig, T., Gorbalenya, A.E., Campanacci, V., Cambillau, C., Canard, B., and Ziebuhr, J. (2006). Discovery of an RNA virus 3“->5” exoribonuclease that is critically involved in coronavirus RNA synthesis. Pnas 103, 5108–5113.
[14]
[15]
[16]
[17]
[18]
参考资料
FIELDS Virology 5ed
(biology)
作者简介
李庆超,微生物学博士,研究生期间于中科院巴斯德所从事病毒学研究,现从事微生物教学和细胞生物学研究。
版权说明:欢迎个人转发,任何形式的媒体或机构未经授权,不得转载和摘编。转载授权请在「返朴」微信公众号内联系后台。
话题:

0
推荐
财新博客版权声明:财新博客所发布文章及图片之版权属博主本人及/或相关权利人所有,未经博主及/或相关权利人单独授权,任何网站、平面媒体不得予以转载。财新网对相关媒体的网站信息内容转载授权并不包括财新博客的文章及图片。博客文章均为作者个人观点,不代表财新网的立场和观点。
 京公网安备 11010502034662号
京公网安备 11010502034662号 {kind=link}