本文归纳了灭活病毒的若干种方式及其灭活原理,大家可据此寻找合适的消杀方法。同时,我们还会看到,新冠灭活疫苗也许还有更多的发展空间。
撰文 | 彭程(清华大学李赛实验室博士研究生)、李赛(清华大学结构生物学高精尖创新中心研究员)
人类的发展历史也是一部病毒抗争史,回顾历史长河,病毒已对人类展开过无数次猛烈的进攻。根据推测,天花病毒在一万多年前就已经出现,直到1980年之前一直在世界各地横行。天花致死率很高,仅在20世纪就造成全世界约3亿人死亡[1]。流感病毒在近一百多年也多次肆虐全球,仅1918年的西班牙大流感便夺去了数以千万计的生命[2]。此外,人类免疫缺陷病毒(俗称“艾滋病毒”)、埃博拉病毒、肝炎病毒依旧对人类虎视眈眈,当前新型冠状病毒也因其突变株的不断出现而持续表现出强劲的传播特性。截至目前,本次新冠疫情的死亡人数也已经上升到人类历史上病毒瘟疫致死人数的第三位(自Wikipedia)。面对这个看不见的敌人,人类仿佛显得有些被动,难道我们不能主动去“杀死”病毒吗?
其实在还未认识病毒时,人类就已经摸石头过河,摸索出了一些“杀死”病毒的方法。我国古代医学对此也有所记载,有些方法现在来看仍具有科学指导意义。如李时珍在《本草纲木》中记载的“天气瘟疫,取出病人衣服于甑上蒸过,则一家不染”就是利用高温“杀死”病毒的例子。随着生物学、物理学、现代医学和化学的进步,人类对病毒的认知也从原来的“邪气”、“上帝的惩罚”到现在的“一种专营寄生的微小生物颗粒”,这也为“杀死”病毒的手段开发提供了生物学依据。
如今,人们已经拥有多种“杀死”病毒的手段,有些能破坏病毒结构而快速“杀死”病毒,如酒精(破坏囊膜)、高温(蛋白变性),这类也是目前消杀的常用手段,有些方法能在“杀死”病毒的基础上保留病毒的免疫原性或结构完整性,如甲醛(固定蛋白)、β-丙内酯(破坏核酸),病毒被它们“杀死”后常被用来制备疫苗或用作病毒的结构生物学研究。
如何才能“杀死”病毒?
其实,“杀死病毒”这个表述并不准确。病毒可谓是大自然以“极简主义”设计的生物体之一,其构造简单到仅剩蛋白/脂膜包裹的核酸。在体外它不会自我复制,没有能量消耗,也没有物质的合成与分解,貌似就是个“死”的状态,但它一旦侵入到细胞内,又可以将整个细胞变成自己的生产工厂,繁殖出更多的病毒颗粒来。因此,病毒是否有生命这个话题一直没有明确的答案。前文所说的“杀死”,其实是指让病毒失去繁殖的能力,也就是让病毒哪怕遇到宿主也别再“活过来”。在体外,这一过程在病毒学界则被称为“灭活”。
若想阻断病毒的繁殖,首先得要了解病毒的生活周期。病毒颗粒通过其表面的蛋白作为“钥匙”识别宿主并进入胞内,利用细胞的原料和能量复制自己的基因组,合成自己的蛋白,这些新合成的基因组和结构蛋白会装配成子代病毒颗粒后被释放到胞外。针对病毒的生命活动过程,灭活的思路大体分为三种,一种是破坏病毒的整体结或病毒的蛋白结构,使其丧失侵入细胞的能力,另一种是“锁住”病毒的蛋白,使其无法发挥功能,最后是破坏病毒的基因组,使其基因组无法在胞内复制。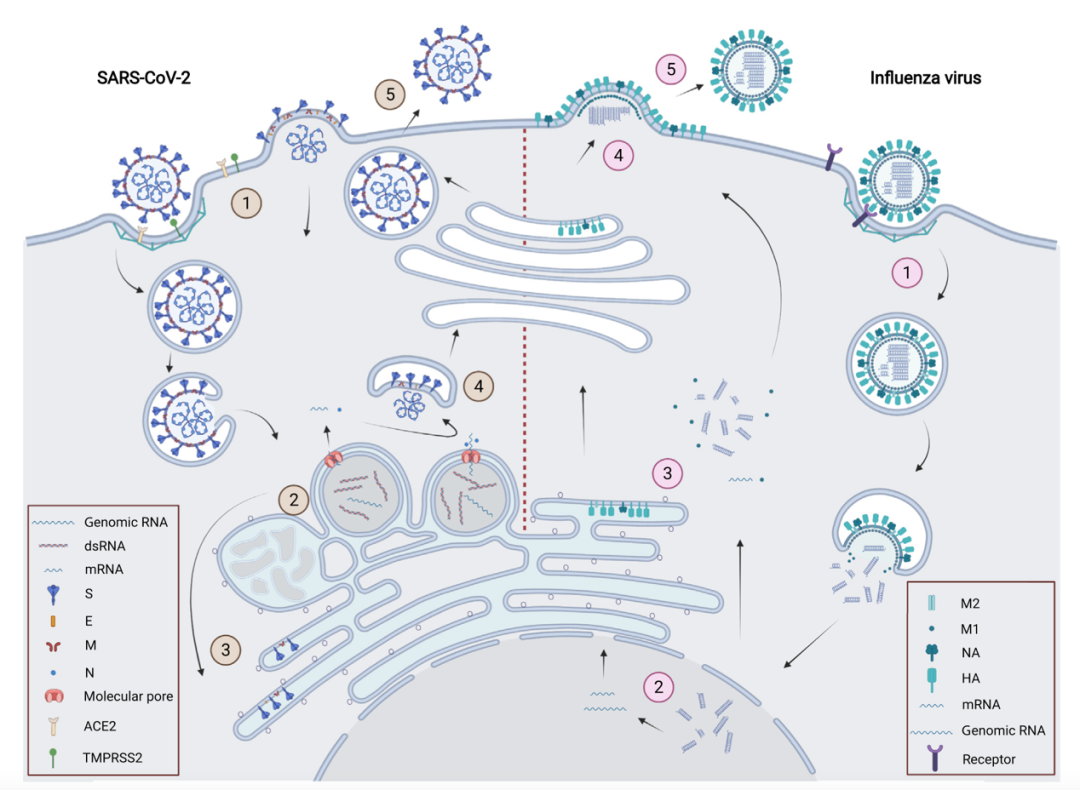
新冠病毒与流感病毒的生活周期[3]。新冠病毒(左)和流感病毒(右)的生命周期都经历以下步骤:①进入细胞。病毒颗粒表面蛋白与受体识别,发生膜融合并释放遗传物质;②基因组复制。新冠病毒诱导细胞产生双膜囊泡(DMV)[4]作为其基因组复制的场所,流感病毒在细胞核中进行基因组复制;③病毒蛋白翻译;④病毒颗粒组装和出芽。新冠病毒的组装发生在内质网-高尔基体中间体(ERGIC),流感病毒在细胞膜上组装;⑤子代病毒释放。
基于以上思路,各种各样病毒灭活的方法也随之诞生。根据灭活方式分类,可分为化学灭活和物理灭活两种。不同的灭活方法对病毒产生不同的影响,有的方法可以快速灭活病毒,有的方法能够在灭活病毒的基础上保留病毒的蛋白结构或免疫原性。科学家们也基于不同的灭活手段进行着下游的科研,如快速消毒手段的建立、病毒的结构解析、灭活疫苗的开发等。本文将简要介绍几种主要的病毒灭活手段及原理。
化学灭活
甲醛
大家对甲醛应该不陌生,它的水溶液就是大名鼎鼎的福尔马林,是一种防腐剂和固定剂。甲醛含有一个电子缺失的中心碳原子,是亲电子的,因此可以与亲核体发生亲核加成反应。它可以使蛋白质氮端的氨基以及侧链含氮的氨基酸(如赖氨酸、精氨酸、酪氨酸等)单羟基化,并进一步脱水形成亚胺中间体,而这些中间体可以与与精氨酸、酪氨酸的残基再次反应形成亚甲基桥,发生交联。此外,甲醛也可以使腺嘌呤单羟基化,阻止基因的读取[5]。因此,甲醛既可以对病毒的蛋白和基因组实现双重打击,还能通过交联固定病毒蛋白的结构。由于甲醛能“锁定”蛋白结构,甲醛灭活既是灭活疫苗开发的主要候选手段,也是结构生物学家在病毒结构解析中常用的病毒灭活方式[6]。
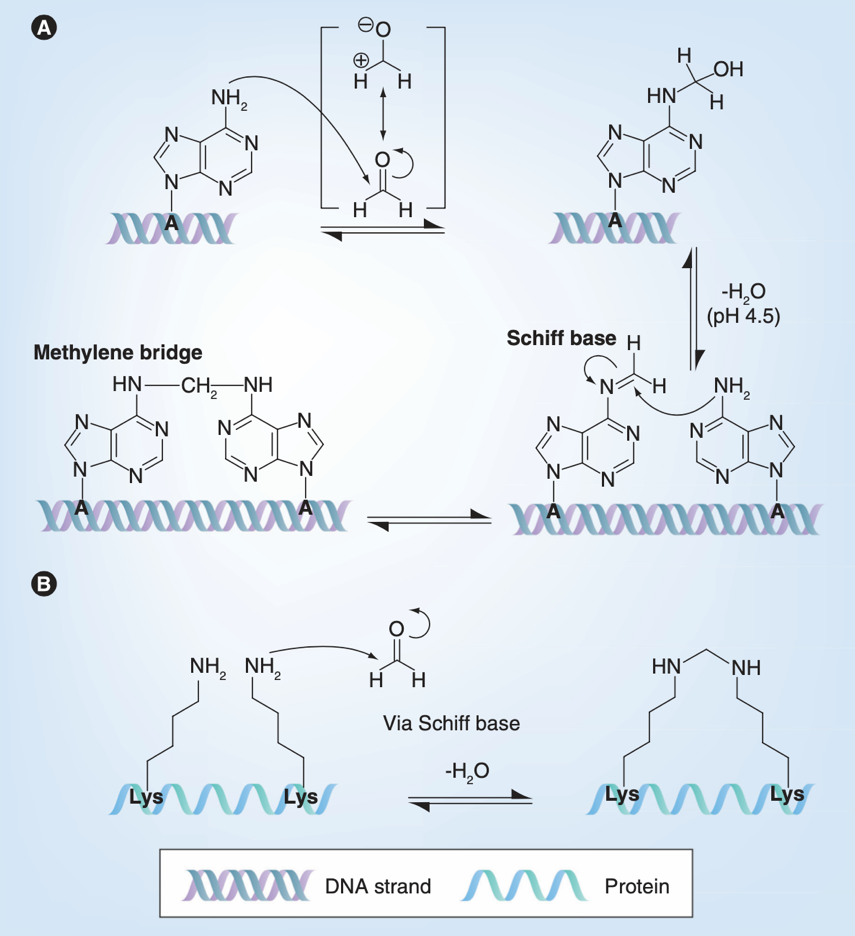
甲醛与腺嘌呤和赖氨酸的反应原理[5]
β-丙内酯
β-丙内酯是一种主要针对鸟嘌呤的烷化剂,也被认为是一种倾向于破坏核酸的灭活试剂。亲电的β-丙内酯可与鸟嘌呤发生亲核取代反应,使β-丙内酯开环和鸟嘌呤烷基化,导致病毒基因组失活[5]。目前,β-丙内酯是灭活疫苗开发中最广泛使用的灭活剂,但有研究表明β-丙内酯也会对部分氨基酸进行修饰,发生酰化和交联反应[7]。
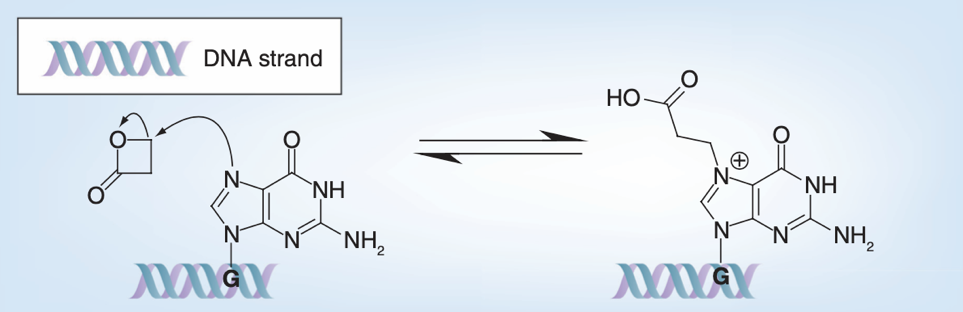
β-丙内酯与鸟嘌呤的反应原理[5]
乙醇
乙醇也就是酒精。乙醇对病毒的灭活速度很快,尤其是囊膜病毒。囊膜病毒是一类外层由脂双层包裹的病毒。由于乙醇具有脂、水双亲性,因此乙醇可以增强膜对水的亲和力,同时减少非极性氨基酸残基之间的相互作用,从而既能破坏病毒的整体结构,又能使病毒蛋白质变性[8]。
2021年,Das等人通过分子动力学模拟的手段描述了乙醇对囊膜病毒的破坏作用,病毒在75%乙醇溶液浸泡后,其囊膜会发生瓦解并导致内容物的释放[9]。新冠病毒也是一种囊膜病毒,2020年Annika Kratzel经过实验发现,新冠病毒在高于30%酒精溶液中浸泡30秒后其感染力可下降至本底水平[10]。除乙醇之外,还有很多醇类化合物也能表现出较快的灭活速度,如正丙醇、异丙醇等[10]。由于这些化合物具有灭活速度快,毒性小的特点,因此常被用作洗手液的主要成分和物品的表面消毒剂等。
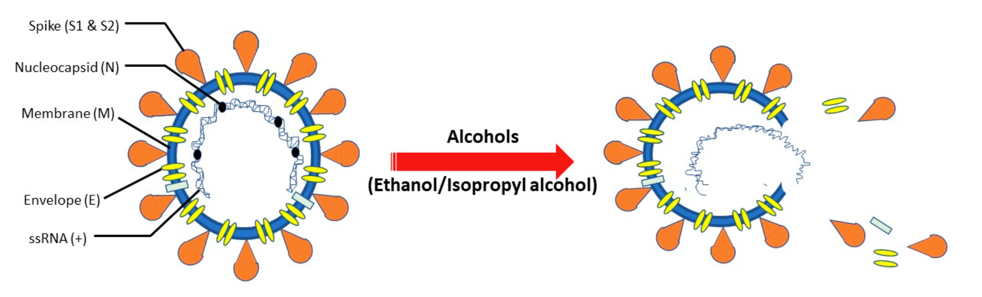
醇类化合物破坏新冠病毒的原理图[8]
物理灭活
温度
高温也是一种常见和常用的灭活手段。在高温的作用下,维持病毒蛋白二级、三级结构的化学键会被破坏,导致蛋白质变性,使病毒丧失侵染细胞和复制的能力。加泰罗尼亚理工大学的Didac Martí等人利用分子动力学模拟发现高温会引起新冠病毒表面的蛋白尤其是表面蛋白的受体结合区域发生结构变化和氢键重排[11]。此外,高温还可以使核酸的骨架发生断裂。
值得一提的是,目前认为“较低”高温(低于41℃)就可以使病毒基因组发生断裂,而不影响蛋白结构和功能[5]。2020年,巴斯德研究院的Christophe Batéjat对新冠病毒的鼻咽拭子样品进行56℃、65℃和95℃的高温处理,发现分别经过了20min、10min和3min便会使新冠病毒失活[12]。高温灭活的手段因其效果稳定且具有广谱性,现在也是作为生物安全废弃物处理的必需步骤。
紫外光
紫外光是频率比蓝紫光高的不可见光。根据国际标准化组织(ISO)的标准,紫外光可根据波长进一步分为UVA波段(320–400 nm)、UVB波段(280-320 nm)和UVC波段(200-280nm)等。其中,UVC对病毒最具有破坏性,随后依次为UVB、UVA[13],因此UVC也是用来灭活病毒的主要波段。
紫外光对病毒核酸的破坏方式主要是诱导核酸内部形成嘧啶二聚体。当紫外光被病毒的核酸吸收后,病毒基因组中相邻或相对的两个嘧啶会发生光化学融合,形成共价连接的嘧啶二聚体。嘧啶二聚体由于共价作用会给基因组的骨架引入张力,导致基因组骨架断裂[5]。对部分病毒而言,紫外光能在病毒完全灭活的基础上保留病毒的融合活性[14]。对于新冠病毒,经过9分钟的UVC照射(累计剂量1048 mJ/cm2)便可使高感染滴度(5 * 106 TCID50/mL)的新冠病毒彻底灭活[13]。虽然UVC的灭活效果好,但其穿透能力较差,普通透明玻璃、衣服、塑料等就能阻挡大部分UVC。因此,UVC常被用在器械物品表面及医院、生物安全等级实验室的室内空间消毒。
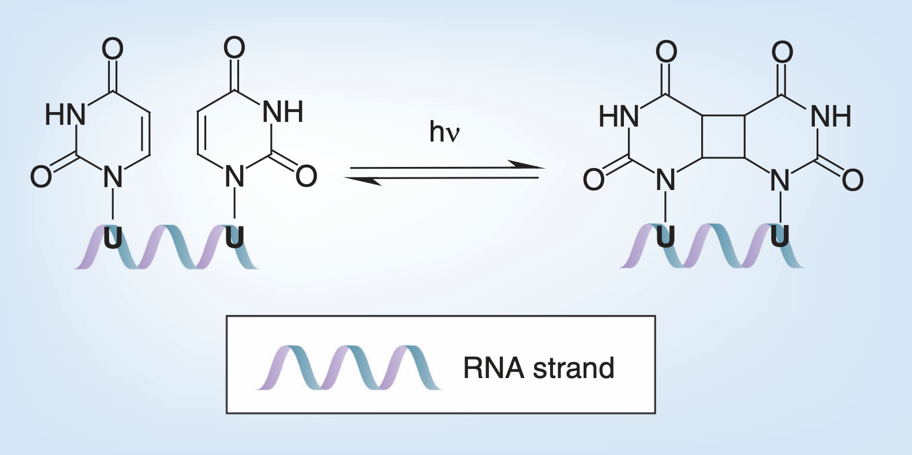
相邻尿嘧啶经紫外光照射形成嘧啶二聚体原理图[5]
电离辐射
电离辐射是一种能导致物质原子或分子发生电离的能量。三种主要的电离辐射技术是γ辐射、电子束和X射线。γ射线是一种由原子衰变裂解时放出的射线。电子束(eBeam)即为从电子束加速器中产生的高能电子束。X射线是由原子核外电子的跃迁或受激等作用产生的射线[15]。虽然来源不同,但是对病毒灭活的机理基本类似。
电离辐射对病毒的破坏作用分为直接作用和间接作用。电离辐射可以直接将病毒体内分子的化学键打断,也可以使水分子发生电离。水分子辐解会产生各种各样的高活性自由基。尽管这些自由基的存在时间极短,但其可以与周围的蛋白和核酸反应,产生极大的破坏性,间接对病毒造成损伤[15]。不过也有人认为电离辐射会主要攻击病毒的基因组而非蛋白质[16]。电离辐射的灭活效果与辐射剂量有关,且不同病毒存在较大差异[16]。随着发射源技术的更新与进步,电离辐射灭活技术也开始应用在了疫苗研发、进口国冷链包装消毒等领域。
灭活的病毒就是灭活疫苗吗?
谈到病毒的灭活,自然就离不开灭活疫苗这个话题。疫苗的作用是为了让我们的机体获得抗原特异的适应性免疫。在佐剂的辅助下,疫苗特定的免疫原性可让我们建立起特异的免疫记忆。当下次相同病原体侵袭时,机体便能快速激活相应的免疫系统,迅速清除病原体[17]。在由疫苗介导的免疫记忆的建立中,疫苗的免疫原性起着关键的作用,灭活疫苗也不例外。
虽然灭活病毒有一定的安全保障,但是并非被灭活的病毒都还具有正确的免疫原性。从分子层面来讲,免疫原性的保留就是病毒表面蛋白完整性和完好性的保留。而从上文可知,病毒灭活的过程中会不可避免地影响蛋白结构。因此,在灭活疫苗研发的过程中需要寻找一个平衡点,即,在保证稳定彻底灭活病毒的基础上,尽可能提高病毒的免疫原性。从上个世纪初开始,科学家便不断尝试用不同的方式制备具有免疫原性的灭活疫苗,其中也有不少已经可以使用和普及的疫苗,包括脊髓灰质炎病毒疫苗、甲型肝炎病毒疫苗和狂犬病疫苗等。
目前,灭活疫苗的灭活方式仍以化学试剂为主,其制备流程大致为:病毒扩增——病毒灭活——化学试剂去毒——病毒纯化——添加佐剂——装瓶。虽然工艺看起来比较传统,但在研发的过程中仍有几大难点亟待解决:
1、在制备疫苗之前,需要获得一株具有良好免疫效应的稳定种子毒株;
2、需要选择合适的灭活剂、剂量及处理方式,以保证疫苗的免疫原性;
3、需要对灭活病毒进行高标准的纯化;
4、需要较高的工业水平以支撑超大规模的快速量产。
这些因素导致一般疫苗从开发到上市都会经历大约10年之久。不过,出于科研、工艺水平的进步和时间的紧迫性,我国在分离出首株新型冠状病毒毒种后不到一年半的时间内,自主研发并上市的灭活疫苗便已经获得了世卫组织的紧急使用授权。
相比于其他疫苗,灭活疫苗具有研发周期短、工艺相对成熟、安全较有保证、易于保存和运输的优势。但灭活疫苗自身也有一些弱点,如为应对病原快速变异的疫情,常需针对变异毒株更新疫苗,核酸类疫苗的更新速度相对灭活疫苗可能更快。此外,对于一些抗原稳定性较差的病毒,传统灭活方式难免会损坏免疫原性。有研究发现,新冠病毒经过0.05%的β-丙内酯4℃敷育36h后,其表面74%的蛋白构象已不再具备正确的免疫原性[18],这也侧面说明灭活疫苗的研制仍有很大的开发潜力。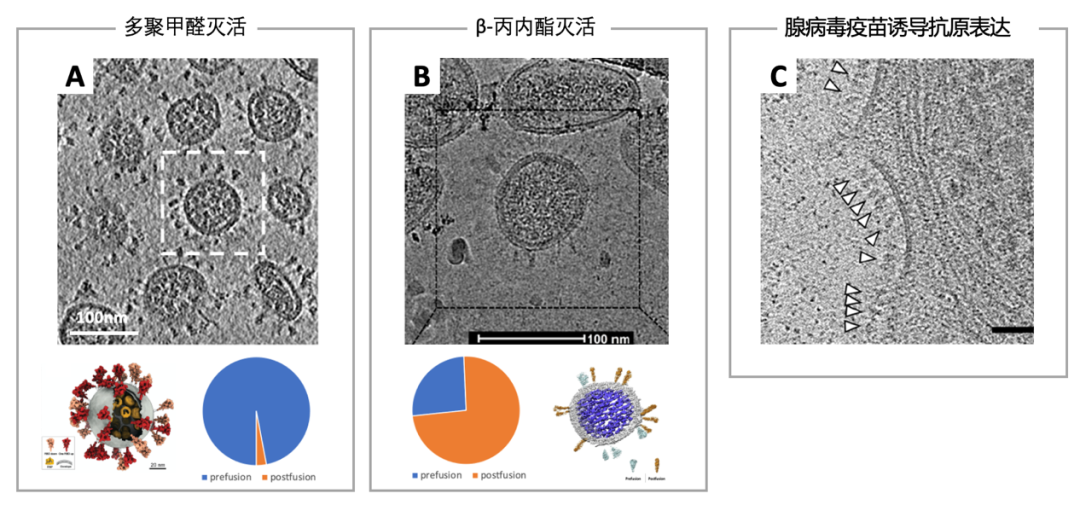
经多聚甲醛[6]与β-丙内酯[18]灭活的新冠病毒的结构和腺病毒疫苗诱导抗原在细胞表面表达的电镜照片[19] 。图A和图B:与多聚甲醛固定后的病毒相比,β-丙内酯灭活可能会造成刺突蛋白发生巨大的构象变化,由倒三角形(饼图中蓝色)变为长柱形(饼图中橘黄色)。但并不排除这剧烈变化是发生在病毒纯化时,因为β-丙内酯处理后的刺突蛋白较甲醛处理后具有更高的自由度。不过,这也并不代表甲醛灭活的病毒免疫原性更高,经研究,经甲醛固定后可能减少了刺突蛋白上受体结合区域的暴露,反而影响了免疫反应的诱导[20]。图C:一类核酸疫苗——腺病毒疫苗可直接诱导宿主细胞表达抗原,刺激机体免疫反应,这种方法引入的抗原不会受到外界损坏。图为细胞接种了腺病毒疫苗后表面表达出大量抗原。
展 望
灭活是我们主动对抗病毒的武器,除直接的消杀外,我们还可以利用灭活的病毒为我们自己添加一堵免疫城墙。寻找仅破坏核酸而不改变抗原结构、成本低、通量高、无毒性的灭活方式,是提升灭活病毒疫苗制备工艺的目标。目前,多种物理灭活的方法也在疫苗开发上表现出很大的潜力,甚至有些方面更有优势。首先,物理灭活不会引入新的毒性,可以减去制备上去毒的操作;另外,物理灭活具有广谱性,对不同种类或能快速变异的病毒适用性更强。此外,物理灭活还有保留较好免疫原性、制备成本低、通量高的研发潜能。目前已有利用电离辐射进行灭活疫苗研发的例子,如用γ射线灭活的流感病毒[15]、脊髓灰质炎病毒[21]等均表现出一定的开发潜能。
随着免疫学理论的不断累积和多种灭活技术的不断成熟,相信在不久的将来,人类在面对疫情的时候可以当即反应,既能迅速消杀,也能快速制备疫苗,主动反击,阻止病毒的扩散和传播。
参考文献
[1] Henderson, D.A., The eradication of smallpox--an overview of the past, present, and future. Vaccine, 2011. 29 Suppl 4: p. D7-9.
[2] Lemon, S.M. and A.A. Mahmoud, The threat of pandemic influenza: are we ready? Biosecur Bioterror, 2005. 3(1): p. 70-3.
[3] Li, S., Cryo-electron tomography of enveloped viruses. Trends Biochem Sci, 2022. 47(2): p. 173-186.
[4] Wolff, G., et al., A molecular pore spans the double membrane of the coronavirus replication organelle. Science, 2020. 369(6509): p. 1395-1398.
[5] Delrue, I., et al., Inactivated virus vaccines from chemistry to prophylaxis: merits, risks and challenges. Expert Rev Vaccines, 2012. 11(6): p. 695-719.
[6] Yao, H., et al., Molecular Architecture of the SARS-CoV-2 Virus. Cell, 2020. 183(3): p. 730-738 e13.
[7] Uittenbogaard, J.P., et al., Reactions of beta-propiolactone with nucleobase analogues, nucleosides, and peptides: implications for the inactivation of viruses. J Biol Chem, 2011. 286(42): p. 36198-214.
[8] Basak, D. and S. Deb, Sensitivity of SARS-CoV-2 towards Alcohols: Potential for Alcohol-Related Toxicity in Humans. Life (Basel), 2021. 11(12).
[9] Das, S., et al., The role of the envelope protein in the stability of a coronavirus model membrane against an ethanolic disinfectant. J Chem Phys, 2021. 154(24): p. 245101.
[10] Kratzel, A., et al., Inactivation of Severe Acute Respiratory Syndrome Coronavirus 2 by WHO-Recommended Hand Rub Formulations and Alcohols. Emerg Infect Dis, 2020. 26(7): p. 1592-1595.
[11] Marti, D., et al., Temperature effect on the SARS-CoV-2: A molecular dynamics study of the spike homotrimeric glycoprotein. Comput Struct Biotechnol J, 2021. 19: p. 1848-1862.
[12] Batejat, C., et al., Heat inactivation of the severe acute respiratory syndrome coronavirus 2. J Biosaf Biosecur, 2021. 3(1): p. 1-3.
[13] Heilingloh, C.S., et al., Susceptibility of SARS-CoV-2 to UV irradiation. Am J Infect Control, 2020. 48(10): p. 1273-1275.
[14] van Duijl-Richter, M.K.S., et al., Chikungunya virus fusion properties elucidated by single-particle and bulk approaches. J Gen Virol, 2015. 96(8): p. 2122-2132.
[15] Bhatia, S.S. and S.D. Pillai, Ionizing Radiation Technologies for Vaccine Development - A Mini Review. Front Immunol, 2022. 13: p. 845514.
[16] Abolaban, F.A. and F.M. Djouider, Gamma irradiation-mediated inactivation of enveloped viruses with conservation of genome integrity: Potential application for SARS-CoV-2 inactivated vaccine development. Open Life Sci, 2021. 16(1): p. 558-570.
[17] Vetter, V., et al., Understanding modern-day vaccines: what you need to know. Ann Med, 2018. 50(2): p. 110-120.
[18] Liu, C., et al., The Architecture of Inactivated SARS-CoV-2 with Postfusion Spikes Revealed by Cryo-EM and Cryo-ET. Structure, 2020. 28(11): p. 1218-1224 e4.
[19] Watanabe, Y., et al., Native-like SARS-CoV-2 Spike Glycoprotein Expressed by ChAdOx1 nCoV-19/AZD1222 Vaccine. ACS Cent Sci, 2021. 7(4): p. 594-602.
[20] Bewley, K.R., et al., Immunological and pathological outcomes of SARS-CoV-2 challenge following formalin-inactivated vaccine in ferrets and rhesus macaques. Sci Adv, 2021. 7(37): p. eabg7996.
[21] Tobin, G.J., et al., A novel gamma radiation-inactivated sabin-based polio vaccine. PLoS One, 2020. 15(1): p. e0228006.

0
推荐
 京公网安备 11010502034662号
京公网安备 11010502034662号 {kind=link}