有些谜团可能永远也无法解开。
撰文 | Amanda Heidt
编译 | 黄炎
“我从哪里来?”
“我从哪里来”,这个问题和“我是谁”“我到哪里去”并列为哲学上的三大基本问题。自有人类文明以来,人们从不曾放弃对自身来源的探索。千百年来,无数文人墨客、才子佳人为之付出了鲜血与生命——好在这种事情只发生在蒙昧时代,现在人们只需要付出时间和头发就可以了。目前,经过几十代人的努力,科学家们基本把这个问题解答到了“我们从原始真核生物演化而来”这一步,但真核细胞的起源仍然扑朔迷离。所以,现在进化学家们要回答的问题是:
“我们从原始真核细胞生物而来,那么原始的真核细胞又从何而来呢?”
2022年,来自巴黎大学萨克雷分校 (University of Paris-Saclay) 的生物学家López-García与同事们一起踏上了继续探究生命起源的旅程。为此,众人前往了世界上气候最为干燥的地方之一——灌木丛生、砾石遍地的南美洲阿塔卡马沙漠北部的高原地带。那里不欢迎访客,但可能拥有关于复杂生命起源的线索。群山和沙丘的包围之下,存活着温暖而苦咸的小水池。氰基细菌和古细菌组成的菌毯,像千层饼一样,一层又一层——对于它们而言,那可能是沙漠之中难得的绿洲。López-García将其称之为“原初之森”,意即早在地球物种大爆发之前便存在的“复杂”生态系统。科学家们现在用这些微小的生态系统来模拟远古的生态系统,因为它们产生的时间肯定不晚于真核生物首次出现。

菌毯可能模仿了早期地球上产生真核生物的条件。DIVERSITY, ECOLOGY AND EVOLUTION OF MICROBES (DEEM)/PURIFICACIÓN LÓPEZ-GARCÍA
这些菌毯的每一层,都有不同种的微生物相濡以沫。占领了光和氧气充足的地表的,主要是蓝藻,它们是最早的光合放氧生物。它们的出现使地球的环境从低氧转向有氧,才有了后来有氧呼吸的物质基础——让我们一起说“谢谢蓝藻”。在这里,蓝藻不仅哺育了人类,下层菌毯中,低氧环境下存在的异养生物以蓝藻的副产物为食。菌毯的更下层垫子又黑又臭,这是缺氧环境下微生物还原硫酸盐、生产甲烷的结果。在这里,大家活得都不容易。互相以代谢废物为食——大家彼此之间一把屎一把尿把对方喂大。
这种一把屎一把尿的关系,科学上称之为“互养共栖”或“营养共生”。López-García表示,这种临时性的互利共生关系可能随着时间的推移逐渐稳定下来,并演化为永久性的关系——约炮约到白头偕老,大概就是这种感觉吧。在这种环境下,不同种微生物的个体可以彼此嵌套在一起,形成一个较为稳定的主体。科学家们怀疑,这种主体就是早期复杂真核细胞的雏形。在漫长的历史中,这种最原始的“细胞”逐渐占据了主流生态位,茁壮成长为稳定的真核细胞,而分工不同的真核细胞又以类似的方式联合起来,最终变成了如今多种多样的宏观生命。这个过程被称之为“真核发生 (eukaryogenesis)”。真核发生的定义尚有争议,但一般指10至20亿年前细胞复杂性激增的演化过程。
这段时间内,细胞核、线粒体、细胞骨架、细胞膜、叶绿体等现代真核细胞的一些标志性特征首次亮相。这些特征出现在所有真核细胞的第一个和最后一个共同祖先之间。按照首字母缩写法,前者被称为FECA (First Eukaryotic Common Ancestor),而后者名叫LECA (Last Eukaryotic Common Ancestor)。但是在此过程中的诸多细节仍然悬而未决。最早的真核生物长什么样?它们可能与其他生物建立何种关系?它们的分子机制是如何运作,又如何进化?……这些谜团有待解开。对于“在哪个生命分支中萌芽”“哪些微生物参与者可能对此做出了贡献”等重要问题,科学家们也尚未达成共识。
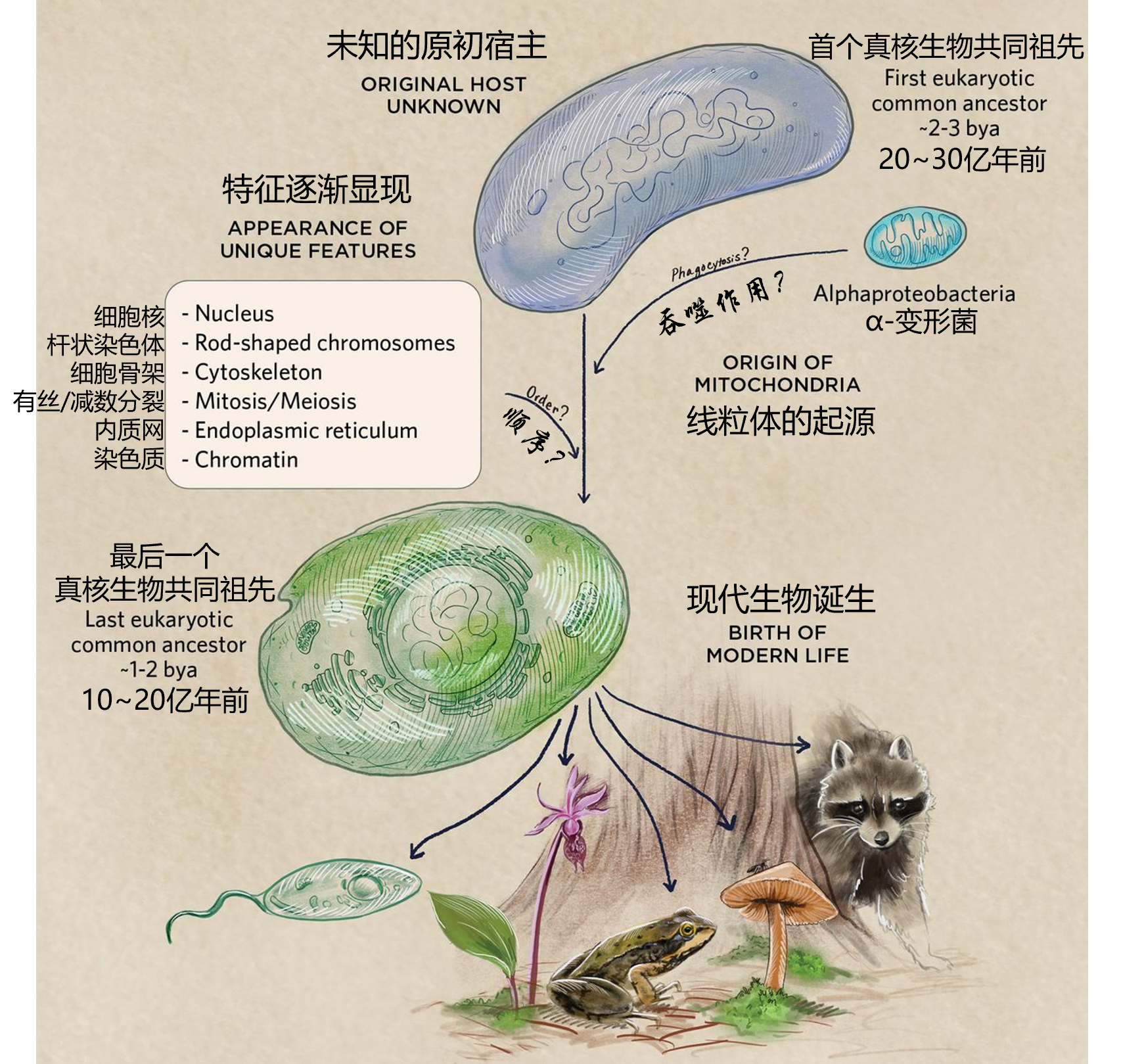
不过,近年来鉴定出的“阿斯加德古菌 (Asgard archaea) ”再一次激活了关于真核发生的讨论。这种取名自北欧神话的古细菌是“现存当代真核生物最近的近亲”,它为过去的讨论提供了证据,又引入了新的问题。一些研究者认为,这是目前生物学上最令人兴奋的发展。“人们发现的是如此之多,而又有如此多的预言正在被验证”。
北欧神宫阿斯加德现世
2013年,拿到进化微生物学博士学位的Anja Spang在寻找一个博后岗位(译者注:谁让你学生物,找不到工作了吧?!)。他的博士毕业课题研究的是一群名为Thaumarchaeota(现已更名为Nitrososphaerota)的古细菌。(译者再注:你看这课题,像能找到工作的吗?!)在博士期间,Anja发现这些古细菌和一些其他古细菌的基因组包含了编码“真核特征蛋白(eukaryotic signature proteins, ESP)”的基因。它们之所以被称为真核特征蛋白,就是因为他们是真核细胞内的特征性蛋白,也就是说它们不应该在古细菌中出现——但它们就是出现了。带着这一疑问,Anja Spang加入了瑞典乌普萨拉大学(Uppsala University)的Thijs Ettema教授课题组,开始了他的深海探秘之旅。(译者本人本科即毕业于瑞典乌普萨拉大学,看到此段时表示:吃瓜吃到自己家.jpg。)

大西洋中部的洛基城堡(Loki's Castle),由五个活跃的热泉喷口组成。(Image Credit: Centre for Geobiology BY R.B. Pedersen)
在挪威和格陵兰之间的北大西洋海面下2300多米处,有一堆被称为“洛基城堡”的海底沉积物,Ettema课题组打算从中提取出基因组。最开始的样本是体积不到一茶勺的海底泥,但是在分析过程中,用来注释和分析遗传物质的软件返回了奇怪的结果——这其实也还好,科研经验表明,意料之外而不尽如人意的结果好就好在它大概率是真的。软件标记了编码肌动蛋白 (actin) 的基因的同源基因。肌动蛋白在真核细胞中用来维持细胞形态,是一种典型的真核特征蛋白,它显然不该在古细菌中出现。所以软件标记的同源基因来自一个新的类群。
2015年,Ettema团队在《自然》杂志上发文,将这一基因命名为Lokiarchaeota,其中的“Loki”即致敬北欧神话中的洛基。随后几年中,该团队逐渐充实了这一类群,并将其命名为“阿斯加德超系(Asgard superphylum)”,除了洛基之外,还包括以雷神、奥丁、海姆达尔等北欧诸神命名的类群——这些神都住在阿斯加德神宫,一家人最要紧系齐齐整整的啦。
此后,研究人员在上述“一家人”中发现了其他真核特征蛋白,比如参与了从泛素信号传导到配子融合等各个生理过程的蛋白质的同源物。真核特征蛋白在这一家子中非常普遍,说明这些微生物可能是与现代真核生物最接近的现存原核生物。并且现代真核生物很可能从古细菌那里继承了其分子运作机制。传统意义上,人们认为现在的真核生物是古代细菌或所谓“原-真核生物”进化而来。但现在这种看法快要被颠覆了。大多数科学家现在认为,现存的真核细胞的共同鼻祖是阿斯加德超系的祖先,或另一群类似的古核生物(古细菌)。
2019年,研究人员首次成功培养了阿斯加德超系的生物,从此能够更深入地研究这家人。他们发现,有一种培养物种个体小、长势缓慢,每两到三周才分裂一次;而另一些微生物可以在短短几分钟或几小时内翻倍。他们将前者命名为 “Candidatus Prometheoarchaeum syntrophicum”(显然这个名字来源于普罗米修斯),而Candidatus Prometheoarchaeum syntrophicum和另一群名叫 “Methanogenium”的古细菌紧密共生。前者通过消化氨基酸和肽获取氮和能量,此过程中产生氢气,氢气又被后者所吸收利用。这一过程可以减少微环境中的氢含量,从而缓解细胞压力。研究阿斯加德超系的科学家认为,这种共生关系可能是真核发生的一个模型。

研究人员于 2015 年从宏基因组数据中首次鉴定出阿斯加德古菌,它被认为是与现代真核生物最接近的现存原核生物。几年后,第一个阿斯加德古菌Candidatus Prometheoarchaeum syntrophicum被培养出来,揭示了其生物学的独特方面。HIROYUKI IMACHI, MASARU K. NOBU, AND JAMSTEC
Anja Spang表示,在其他古细菌的基因层面也检测到了这一共生关系,但是Candidatus Prometheoarchaeum syntrophicum和Methanogenium的关系为此提供了坚实的证据。多年以后,已经成功上岸并在荷兰皇家海洋研究所 (Royal Netherlands Institute for Sea Research) 建立了自己课题组的Anja回忆说:“当我第一次得知描述这种生物及其营养共生关系的论文终于能发表时,我打心眼儿里高兴。这证明了此类试验工作对于阿斯加德超系的代谢预测是有意义的。”
众说纷纭的真核发生假说
这些早期的观察结果引来了大量的研究——和灌水。在随后的几年里,bioRxiv上涌现出数百篇有关阿斯加德超系和真核发生的论文预印本。
前述发现最直接的影响是将真核生物和古细菌划入了同一个域,从而将真核生物、原核生物和古细菌组成的三域生命树模型缩减为了双域模型。(译者注:“域”是生物分类学中的最大单位,一个域下设有若干个界,随后即是熟悉的门纲目科属种;例如人类即属于真核生物域、动物界、脊索动物门、哺乳纲、灵长目、人科、人属、智人种……)
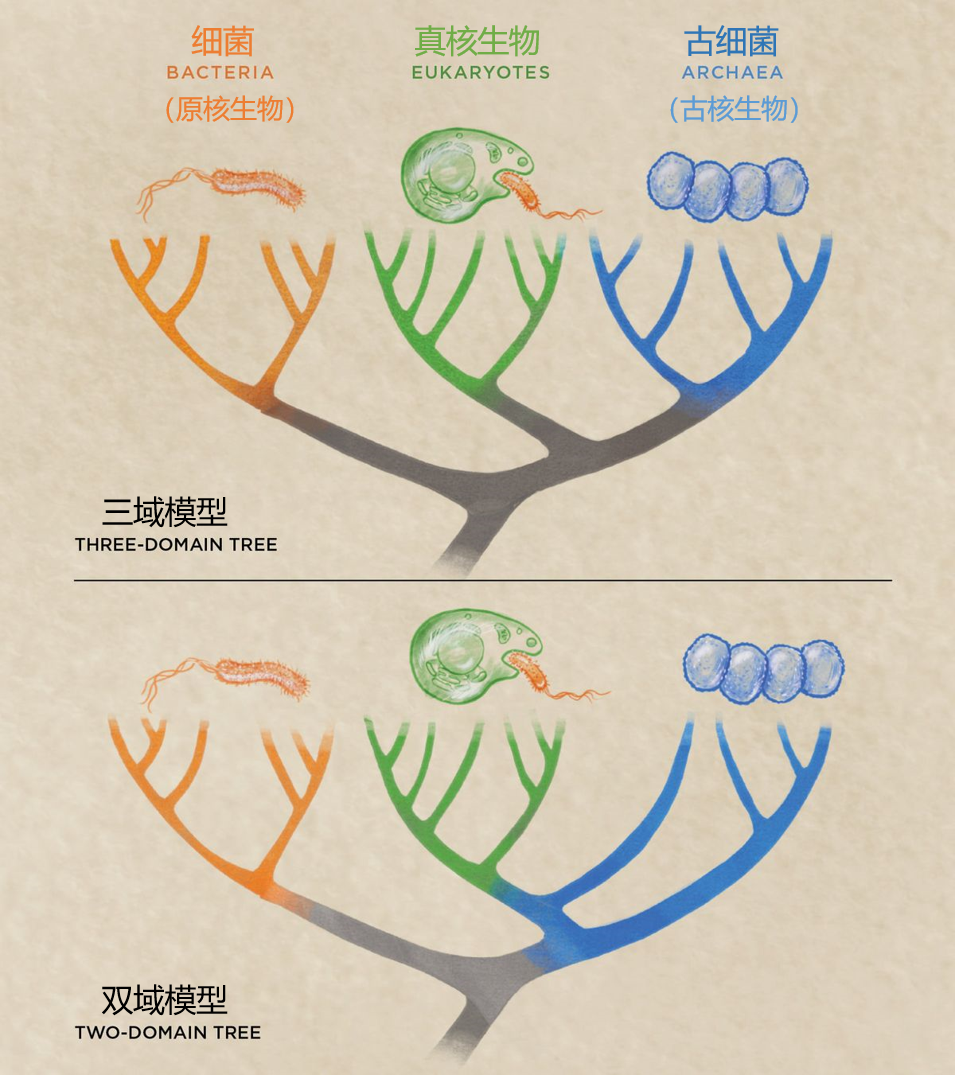
曾经人们认为细菌和古菌都是原核生物,但计划生物学的不断研究,人们发现古细菌和其他原核生物的区别越来越大,于是把古细菌独立出来归为古核生物。
在传统的三域模型中,真核生物和古细菌虽然有共同先祖,但却属于不同的分支。近年来系统生物学领域的研究表明,复杂的真核细胞来源于古细菌。这使得真核生物和古细菌可以被划分为同一个域。在阿斯加德超系被发现之前,人们就已经在争论双域系统,但阿斯加德超系为其提供了更多的证据。双域假说也支持真核生物来自古细菌,而不是所谓的‘原-真核生物’”这一理论。
01
先有细胞还是先有线粒体?
许多科学家都相信,第一批真核生物是从古细菌和细菌之间的合作中进化而来的。在这一过程中,细菌以某种方式进入古细菌内部,并成为了细胞核、线粒体等细胞器——这是真核生物的决定性标志。此过程中的细节仍然模糊不清,但线索最有可能来自于线粒体。“线粒体中存在DNA,这些DNA最早可以追溯到α-变形菌 (alphaproteobacteria)。”法国国家科学研究中心(France’s National Centre for Scientific Research, CNRS)的进化微生物学家Laura Eme表示,“即便我们不知道这一过程的确切路线,但这已经算是实锤了。”
α-变形菌是变形菌门(Proteobacteria)下的一个纲。其内部差异极大,相似之处极少,俗称“散装纲”。α-变形菌大多都呈革兰氏阴性,典型成员有根瘤菌等植物共生菌、沃尔巴克氏体等内共生细菌,以及立克次体等胞内寄生菌。有人认为α-变形菌处在转化为细胞器的路上,而目前的细胞器则是已经上了岸、在胞内有了“编制”的α-变形菌。
八卦即使有了实锤,吃瓜群众也要扒一扒细节——科学家也一样。关于α-变形菌在细胞内“考编上岸”的这一过程,一直存在着不同甚至彼此对立的假说。α-变形菌是怎么进入细胞里的?科学家们咂摸了半天,也没搞明白:胞吞过程需要消耗巨大的能量,对于这么一种堪称奢侈的生理功能,究竟是先有了线粒体为这一过程供能,细胞才演化出胞吞功能,还是先有了胞吞功能,才能把线粒体吞进细胞内?所以科学家们分成 “先有线粒体”和“先有胞吞”两派,在学术期刊上展开了唇枪舌剑。无论如何,长久以来在原核生物身上是没有观察到胞吞这一功能的。
直到最近,研究人员在一种细菌身上发现了一种“类胞吞作用”。Laura Eme对此评价道:“很多人觉得从前原核生物不能胞吞,所以线粒体的祖宗不可能是被吞进细胞的。但是现在,至少我们知道原核生物可以胞吞了。”
生物和化学这种学科折磨人的地方就在这里——和数学、物理等简洁明快的公式化学科不同,化学和生物总是充满了例外。打个比方说,数学和物理对于“北京人”的描述可能是“身份证号110开头的是北京人”,不管这个说法对不对,好歹有一个明确的判断标准。而化学和生物的说法则更接近于“爱喝豆汁儿的人是北京人,但是我们发现了有些人确实在北京出生却不爱喝豆汁儿,所以我们将其定义为‘广义的北京人’,而将传统上定义的北京人称为‘经典北京人’;现在我们有了‘超北京人域’,其下可以分为‘经典北京人’和‘非经典北京人的广义北京人’两界;随后我们又发现了一些人也爱喝豆汁儿但却不是北京人,所以我们将其称之为‘拟北京人’……”
此外,对阿斯加德超系的初步观察还发现了一种另外的吞噬机制。当科学家第一次培养Candidatus Prometheoarchaeum syntrophicum时,他们注意到了胞体表面的一些细长突出部——被称之为“泡”的膜结构的延伸。在上文所述的肌动蛋白同源物的协助下,这些气泡可能能够包围一个外部实体并融合在一起,从而将外物包在里面。如此一来,有关吞噬作用的难题“越来越不是事儿了”。也就是说,α-变形菌很有可能是被“吞”进原核生物里变成线粒体的。
02
细胞核是怎么出现的?
但是,当谈到细胞核时,事情就不那么清楚了。一般认为,真核生物有别于原核生物的最典型特征是“有细胞核”。而有关细胞核生成的猜想范围很广,从“变形虫体内寄宿的细菌”到“远古巨型病毒的残留”,不一而足。上世纪90年代,López-García针对真核生物的起源提出了“营养共生假说”,假设两种细菌与一种古细菌之间存在共生关系。
阿斯加德超系被发现后,López-García和她的同事几年前更新了这一假说。他们并未将古细菌作为原始宿主,而是提出了一个“始祖菌”的概念。在他们的假说中,“始祖菌”是一种与阿斯加德超系中生物类似的、能够产生氢气的古细菌,也是最初的细胞核。而接受这样一颗“细胞核”的宿主大概率是一类δ-变形菌,也即线粒体和α-变形菌的祖宗。现代真核生物中的大多数基因实际上源自细菌,而不是古细菌;组成真核细胞细胞膜的脂质在结构和成分上更接近于细菌而非古细菌(这一现象称为“脂质分离”(lipid divide))——这些事实都能支持他们的假说。而他们的假说也是迄今为止唯一一个既能解释细胞核的起源,又能解释脂质分离现象的假说。
Michelle Leger是巴塞罗那进化生物学研究所 (Institute of Evolutionary Biology in Barcelona) 进化微生物学领域的一位博士后,他的主要工作是对现存古细菌基因组进行溯源,用以支持或反对目前的各类假说。对于López-García等人提出的“营养共生假说”,Leger表示,线粒体中有清晰的α-变形菌的基因组信号,但他还没有发现细胞核中有δ-变形菌的类似信号。
Leger也认为目前的证据确实支持“细胞核的起源是古细菌”这一论断。虽然古细菌的基因只在现在的核基因组中占一小部分,但是这些基因很多都高度保守,例如负责DNA复制和转录的基因很大程度上来自于古细菌。因此,Leger相信“这些假说是有意义的,只是我们至今尚不清楚还有哪些生命体参与了这些进化过程。”
解读真核发生的新途径
很多科研人员表示,纵然已测序到的细菌和古细菌物种数正在飞速增加,为厘清此类生物与早期真核发生的关系提供了新的线索,但令人遗憾的是,此领域的很多问题可能永远不能得到完美的解答了。
真核生物已经出现了太久太久,太多的基因在太多的物种之间交换、传递。科学家不可能将所有东西拼凑在一起,但他们依然在尝试。目前学术界广为采用的研究方法包括组学、分子生物学、化石研究等。
下一个灌水的好地方将会是现代真核生物基因组和蛋白质组的功能研究。研究了功能,就可以提示单个基因和蛋白质在早期祖先中的行为。几年前,只有一个阿斯加德超系基因组,但现在已经有数百个这样的群体,研究人员正在发掘它们的细节。“现在我们清楚地知道真核生物中的哪些基因是从阿斯加德古细菌那里遗传的,这里有很多新奇的东西,”Laura Eme说,“但我们不知道的是,这些基因曾经在阿斯加德超系中做了什么,又正在做什么——这也正是关窍所在。
还记得前面提到的,古菌中有编码真核肌动蛋白基因的疑似同源基因吗?2020年,研究人员合成了阿斯加德超系基因组中的这类同源基因。他们将这些同源物注射到兔细胞中,发现它们可以与真核肌动蛋白结合并执行类似的功能,例如辅助钙离子跨膜。这说明,在真核生物出现之前,钙控制的肌动蛋白细胞骨架可能就存在于古细菌中。
除了培养古菌来研究功能,还有人直接研究“小微化石”。所谓小微化石,就是岩石中早期细胞的显微印迹。加州大学圣塔芭芭拉分校 (University of California, Santa Barbara) 的古生物学家Susannah Porter就认为化石研究同样可能揭示真核发生的途径。她说,当宏基因组测序出现时,化石似乎失宠了,但许多系统发育树依赖于一种叫做“分子钟”(molecular clock)的方法,即,用化石来锚定分析时间。此外,化石本身可能也有用处,它们能够帮助科学家确定某些外部特征在何时首次出现。而Porter正在研究的标本就可以确定早期真核生物进化过程中一些事件的发生顺序。“我们确实收藏有过去20亿到10亿年的化石,它们并没被物尽其用。也许我们可以利用化石的这些特征来拼凑真核细胞的成因。”总之,目前来看,虽然基因组学和分子生物学已经发展到了一定高度,但传统的化石证据依然在微生物进化学和真核发生领域的研究起着重要的作用。
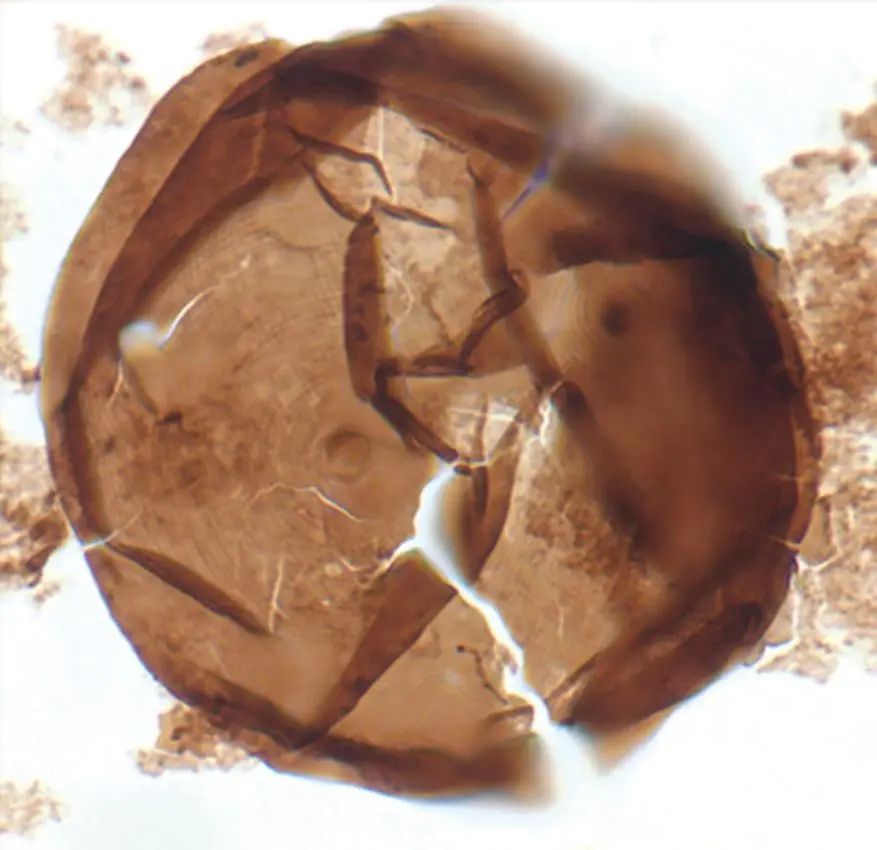
7.5亿年历史的微化石 Valeria lophostriata
COURTESY OF SUSSANAH PORTER
为了避免“孤证不立”,科学家们也在寻找别的证据,用来为化石证据提供佐证。例如,荷兰乌得勒支大学 (Utrecht University) 的计算生物学家Berend Snel最近使用了一种名叫“基因重复段落”的方法进行研究。这种方法假设系统发育树(phylogenetic tree)上的各个分支的长度与其发育时间相关。然而这一方法备受争议,以至于Snel本人都不得不承认其可能存在缺陷。但Snel也表示,进化是一个连续的过程,当且仅当人们将这一漫长过程中的一个个小片段连缀起来之后,真实而完整的进化图谱才会展示在人们面前。
Michelle Leger也同意的一点是,现阶段人类对真核发生的理解还如同婴儿对世界的理解一般。“这些深层进化问题的部分性质是,我们永远不会知道,也永远不会有一个明确的证据来证明我们的假设,但这并不妨碍我们继续完善我们的想法。
另类假说:病毒才是祖先
关于细胞核起源的很多内容都是推测性的。有一种假说认为,现代真核生物的细胞核可能来自原核宿主和病毒之间的合作关系。
在21世纪之初,日本名古屋大学医学院 (Nagoya University School of Medicine in Japan) 的分子生物学家Masaharu Takemura注意到,一组病毒(痘病毒)的DNA聚合酶与真核生物中发现的DNA聚合酶非常相似,并且痘病毒通过创建“独立隔间”在宿主细胞内复制。与此同时,生物技术公司MicroBioGen的研发主管Philip Bell同样对真核生物和细菌之间的差异感到困惑。例如,真核染色体是线性的,而细菌染色体是环形的。细胞核的许多特征并不支持细菌起源。这两名研究人员都在2001年前后脚发表了各自的论文。并且这两个课题组在得知阿斯加德超系及其研究成果之后,都更新了自己的病毒起源假说。
此后,研究人员鉴定出了2003年首次发现的巨型病毒,这些病毒比大多数病毒大得多,具有足够大的基因组,且组内有与各种代谢过程相关的基因。现在,Masaharu Takemura、Philip Bell等人认为这种巨型病毒可能是最初的细胞核。巨型病毒在复杂的隔室中复制,这些隔室看起来与现代细胞核非常相似。它们都很大,也都包含内膜和外膜,并且还都携带了编码宿主细胞运行所需的蛋白质的基因。
然而,“细胞核可能来自一种病毒”的观点一直很难推销出去。它缺乏结构性证据,也缺乏现有数据的支持。不过,研究早期原核生物代谢的微生物学家Valerie De Anda并没有因为“病毒假说”目前缺乏证据支持而被吓退。她和她的同事们目前正在寻找参与转录和翻译的mRNA封端基因。他们认为这些基因来自很久以前的“第一个真核生物细胞核的祖先”。
当谈到自己的理论不被科学界所普遍承认时,Valerie De Anda不免有些懊恼。这使她不由得想起了自己上学的时候老师教过的一首中国的古诗:
自小刺头深草里,
而今渐觉出蓬蒿;
时人不识凌云木,
直待凌云始道高。
参考文献
[1] Shiratori, T., Suzuki, S., Kakizawa, Y. et al. Phagocytosis-like cell engulfment by a planctomycete bacterium. Nat Commun 10, 5529 (2019). https://doi.org/10.1038/s41467-019-13499-2
[2] Martin William F., Garg Sriram and Zimorski Verena (2015) Endosymbiotic theories for eukaryote originPhil. Trans. R. Soc. B3702014033020140330 http://doi.org/10.1098/rstb.2014.0330
[3] Moreira, D., López-García, P. Symbiosis Between Methanogenic Archaea and δ-Proteobacteria as the Origin of Eukaryotes: The Syntrophic Hypothesis. J Mol Evol 47, 517–530 (1998). https://doi.org/10.1007/PL00006408
[4] Caforio, Antonella et al. “Converting Escherichia coli into an archaebacterium with a hybrid heterochiral membrane.” Proceedings of the National Academy of Sciences of the United States of America vol. 115,14 (2018): 3704-3709. doi:10.1073/pnas.1721604115
[5] Vosseberg, J., van Hooff, J.J.E., Marcet-Houben, M. et al. Timing the origin of eukaryotic cellular complexity with ancient duplications. Nat Ecol Evol 5, 92–100 (2021). https://doi.org/10.1038/s41559-020-01320-z
[6] Chaikeeratisak, Vorrapon et al. “Assembly of a nucleus-like structure during viral replication in bacteria.” Science (New York, N.Y.) vol. 355,6321 (2017): 194-197. doi:10.1126/science.aal2130
[7] Mills, Daniel B et al. “Eukaryogenesis and oxygen in Earth history.” Nature ecology & evolution vol. 6,5 (2022): 520-532. doi:10.1038/s41559-022-01733-y
本文经授权编译自:https://www.the-scientist.com/features/the-long-and-winding-road-to-eukaryotic-cells-70556?_ga=2.200353838.1403010427.1666753497-1684915652.1666340439

0
推荐
 京公网安备 11010502034662号
京公网安备 11010502034662号 {kind=link}