阅读:0
听报道
撰文 | 刘文豪
2010年,美国Scripps研究所的Ardem Patapoutian课题组在《科学》杂志报道了哺乳动物中首类介导阳离子内流的机械门控离子通道——Piezo蛋白家族[1],平地惊雷般的开启了一个领域。因与触觉等关键的基础机械感觉关系密切,该蛋白自发现伊始便作为明星分子活跃在各大学术期刊上。Ardem更是因这一发现被授予2021年诺贝尔生理学或医学奖!笔者的导师肖百龙教授曾是Ardem的博后,他见证和参与了Piezo蛋白早期研究的关键工作,独立后在这一领域继续深耕,因而我有幸进入到这一热火朝天的领域中。Piezo蛋白究竟是什么?它们是如何工作的?它们的重要性在哪儿?带着这些问题,让我们一起进入Piezo的世界,一窥科学家们逐步认识Piezo蛋白的过程。
图1. Ardem Patapoutian因其在触觉感知领域的重要贡献获得2021年诺贝尔生理学或医学奖丨来自诺贝尔奖委员会官网
我们对这个世界最初的感知通常是从触觉开始的。第一声啼哭之前,婴儿经历过诱发哭声的拍打;来到人世,感受亲人的爱抚和拥抱;儿时抓周一通乱摸,成长中提笔、拍球、弹拨乐器;日常生活中握手,敲击鼠标键盘,刷手机……触觉充斥着我们日常生活的方方面面。
触觉是躯体机械感觉的一种,除了触觉之外,还有机械痛(区别于日常的轻触觉)、本体感觉(对四肢移动和体位的感知)等也属于躯体机械感觉。这些机械感觉和视觉、嗅觉、味觉和听觉构成了我们日常意义上的五感(听觉也是感知声波振动的一种机械感觉)。

图2. 五种感觉丨来自Bing
这些基本感觉从形成、传递到感知,生物学过程大体上是类似的:特定的感受器官和细胞上特定的受体接收外界刺激,产生动作电位,这种电信号沿神经突触向中枢神经系统传递,最后到达大脑特定区域,就产生了特定的感觉。在这一过程中,作为最初级受体的一系列蛋白质是十分神奇的存在。
人体的蛋白质都是由20种基本的氨基酸组成。可就是仅仅凭借这20种基本组件的排列组合,生物居然能演化出如此精准强大的、感知外界各种物理化学信号的接收器——受体蛋白。以人类为例,眼睛中的光敏感受体可响应380-780nm波长的电磁波;鼻子中的嗅觉受体可帮助我们分辨出空气中的数万种小分子;味蕾上的味觉受体可分辨苦咸酸甜鲜;人耳中声波的受体可以分辨出各种不同响度、频率和音色的声音;遍布全身的触觉受体能帮我们精确分辨出压力、振动、纹理等多种不同的触觉感受。而在动物中,这些受体的功能往往比人类更加强大。因此,鉴定出这些受体究竟由何种蛋白构成,一直都是神经科学家们孜孜以求的关键目标。
视觉上的研究首开先河,视网膜上的感光蛋白(视紫红质)为德国科学家在19世纪发现,而后在上世纪80年代确定了全长DNA序列[2, 3]。接着,仿照研究视觉受体的办法,科学家们相继鉴定到嗅觉和味觉受体的分子基础[4, 5],其中嗅觉受体的工作获得了2004年诺贝尔奖。这三种感觉的受体蛋白或在相应感官中高表达,或在基因序列上有高同源性(三者均属于G蛋白偶联受体家族),这两个特点为它们的鉴定提供了条件。
至于余下的两种机械感觉,造物主似乎给我们出了难题:触觉和听觉的受体蛋白的低表达给生化鉴定带来困难,且在基因序列上也不像视味嗅觉般有章可循[6]。凭借遗传筛选,科学家得以在较低等的生物中鉴定到部分机械感觉通道(细菌中的机械通道MscL倒是生化方法获得的[7])。
而在哺乳动物中,这个疑问一直持续到2010年。这一年,Ardem课题组鉴定到了哺乳动物中首类介导阳离子内流的机械门控Piezo阳离子通道[1],解决了谜题。这一里程碑式的发现为触觉受体的分子基础提供了可能的解释。后续的研究发现,Piezo家族中的Piezo2确实参与触觉感知。
如此重要的一类蛋白是如何千呼万唤始出来的?它的功能是怎样被一步步揭示出来的?今天,我们将从分子水平来讲述Piezo这栋大厦的地基是如何打下的。
在当代生物学研究中,一个好的工作,往往都需要由浅入深的逐步上升到分子水平。具体来说,需要揭示这一过程的各个部分:一个生物学现象的组织器官基础,与之相关的细胞,细胞上行使功能的蛋白,以及编码该蛋白的基因。蛋白质和基因这种分子水平的研究是一个领域的基石。
电光火石——机械力能直接打开离子通道
哺乳动物细胞能产生数万种蛋白,如何从中发现机械敏感蛋白?我们不妨先把问题转化成:哪些蛋白可能响应机械力?
机械力势必引发细胞形态的改变,因此,和细胞膜有密切关系的蛋白都能成为怀疑对象,包括直接定位在细胞膜上的膜蛋白、支持细胞结构的细胞骨架蛋白以及细胞外基质等。
1979年,研究人员用电生理的方法,在牛蛙的听觉上皮细胞记录到了机械力刺激下产生的电流[8]。由于刺激后电流产生的时间在亚毫秒级别(40μs),这个电光火石的瞬间不足以让细胞内第二信使完成信号传递(上述的视觉嗅觉味觉的G蛋白偶联受体就需要借助第二信使产生电流),所以只能是目的蛋白受到机械力后直接产生了电流。这个潜在的机械敏感蛋白,最可能是离子通道[8, 9]。
离子通道是膜蛋白的一种,一般由跨越细胞膜内外的孔道区域和其他辅助元件组成。在受到适当刺激时,孔道区域可打开,依据离子通道对不同离子的选择性和细胞膜内外不同离子的电化学梯度,使部分离子穿越细胞膜,带来细胞膜电位的变化,或由离子自身参与信号传递而形成生物学效应。
无独有偶,1984年,研究人员在鸡的骨骼肌细胞中记录到了由牵拉所致的机械敏感电流,该电流也被认为由离子通道介导[10]。
搜寻范围由此大大缩小——离子通道中就存在着机械敏感蛋白。
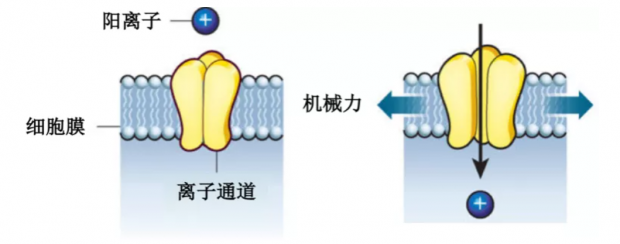
图3. 机械力驱动离子通道打开完成阳离子内流丨改编自Hoffman, et al., 2011, Nature[11]
上下求索——你究竟是谁?
和G蛋白偶联受体家族不同,不同的离子通道在序列和功能上彼此之间差异巨大,唯一被认可的共性是:都含有两次以上的跨膜区[1](膜蛋白都需要插入细胞膜的跨膜区,来定位在细胞膜上,最常见的跨膜区是α螺旋)。
如何判定一个通道是机械门控离子通道呢?科学家们制定了以下几条准则:该通道要能直接被机械力激活而无需通过中间信使;该通道参与机体机械传导的过程;在非机械敏感细胞中表达该通道能赋予该细胞机械敏感性;该蛋白有成孔区和机械敏感区[12, 13]。
此等严苛的标准,加上机械敏感通道常常低表达的特性,使得常规生化手段束手无策,仅在高表达的细菌机械敏感通道MscL的鉴定中方有用武之地[7]。而后世找寻其他真核生物的机械通道大都经由遗传筛选得到。遗传筛选是基于中心法则——蛋白质是由基因编码产生。在个体或者细胞中,让感兴趣的基因发生变异,从而导致该基因对应的、行使生物学功能的蛋白质发生变异或不再表达,如果此时机械感觉相关的表现异常,那么该基因就很能作为备选基因。
在易于进行遗传操作的低等物种中,遗传筛选帮助我们确认了部分机械门控离子通道,比如线虫里的DEG/ENaC通道[14, 15]、果蝇的NOMPC通道[16]。而在脊椎动物,尤其是哺乳动物中,要确定机械敏感蛋白,可以寻找这些低等物种机械通道的同源蛋白。但这种方法收效甚微:哺乳动物中或是不存在相关的同源基因,而有同源基因的,其表达的蛋白却没有机械敏感性。此前唯一确定的哺乳动物机械敏感蛋白——K2P家族的作用是使细胞超极化而变得更加“平静”[17],可是触觉、听觉过程需要让细胞去极化而“兴奋”起来。
那么,我们苦苦找寻的这种受体究竟是什么呢?
学界路漫漫,上下而求索。
平地惊雷——Piezo蛋白横空出世
故事开始于十多年前。彼时,Ardem实验室的博士后Bertrand Coste接下找寻哺乳动物机械门控离子通道这一极具挑战的课题。此前他们已经知道一种鼠源神经母细胞瘤细胞系(N2A细胞)是有较强机械敏感性的:玻璃尖头戳细胞的力刺激和抽吸细胞膜的力刺激均能引发稳定电流。他们试图从这种细胞出发,寻找那个隐藏在重重迷雾里的机械敏感通道。
细胞系易于在体外大量培养,且细胞之间一致性很高,这样的特点允许我们鉴定到细胞内高表达的基因。借助基因芯片,N2A细胞系内高表达的基因被确定。再结合离子通道“至少有两次跨膜”这一特点,他们挑选出一系列备选基因。进一步的,他们用RNA干扰技术逐个抑制备选基因表达,如果某个基因抑制后的N2A细胞失去机械敏感性,那么该基因表达的蛋白极有可能就是那个受体。
事情就这样成了!在Bertrand口中的那个“美妙的时刻”,列表中的第72个基因被抑制后,N2A的机械敏感电流显著下降!这个原名Fam38A的基因后来被命名为Piezo1,同一家族的另一个蛋白Fam38B被命名为Piezo2。Piezo一词起源于希腊语,有“压力、挤压”的意思,用它做名字,寓意该基因是压力受体。
进一步的,他们将Piezo表达在几无机械敏感性的HEK293T细胞系中,成功赋予其机械敏感性。同时,又在哺乳动物传导机械感觉的背根神经节中,发现了Piezo2的高表达,提示了Piezo2和触觉、痛觉感知的密切关系[1]。
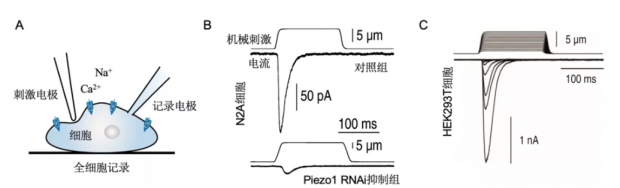
图4. 机械敏感蛋白Piezo1的鉴定
A,用“戳”的机械力刺激细胞的电生理实验设置。记录电极与细胞形成全细胞记录模式,玻璃刺激电极在电驱动下戳细胞。B,上图,在N2A细胞中,戳细胞可诱发内向电流。下图,用RNAi技术抑制Piezo1表达后,N2A细胞的机械敏感电流大幅降低。C,过表达Piezo1的HEK293T细胞展示出显著的机械敏感性。(图B, C改编自Coste, et al., 2010, Science[1])
这是一件平地惊雷的大事!从此哺乳动物机械力受体研究的历史焕然一新!Piezo蛋白是哺乳动物细胞具有机械敏感性的充分必要条件!
然而,这个研究并不是完美的,它仅证明到Piezo是机械门控阳离子通道至关重要的部分。
那么,Piezo蛋白就是那个通道吗?
生化艺术——真的是通道
上述研究虽然证明了Piezo蛋白对于机械敏感电流的出现不可或缺,但不能排除一种可能:Piezo只是这个机械敏感通道的一部分,仅有辅助和调节功能。那么,Piezo本身就足以实现通透离子的通道功能吗?或者说,Piezo蛋白就是这个机械敏感阳离子通道的通道区吗?
一个证据来自果蝇。研究发现,果蝇Piezo和鼠的Piezo在一些和孔道直接相关的性质上有较大差异,比如通道的电导和抑制剂作用能力很不相同。这说明,Piezo的来源不同,孔道相关的电生理特性不同,这暗示着Piezo本身就是通道[18]。
而更直接的方法是:直接提取纯的Piezo蛋白,并重组到人造的类似细胞膜的脂质体上。如果仍能记录到电流,那便证据确凿。
这就要求我们能提取出保持生理功能的Piezo蛋白——这个步骤称为“蛋白纯化”。
蛋白的纯化需要依据每种蛋白的特点量身定制,膜蛋白提取更是其中很棘手的一类。Piezo是很大的蛋白:超过2500个氨基酸,预测有30次以上的跨膜[1]。并且,离子通道经常以多聚体形式发挥功能,使提纯难度进一步增加。因此,Piezo蛋白的纯化非生化功力深厚者不可为之。
彼时,Ardem实验室的另一个博士后,肖百龙博士,凭借其扎实的生化技术,解决了这一难题。纯化的鼠源Piezo1重组入人工脂质体后,确实记录到了通道开放产生的电流,且该电流能被钌红(一种广谱阳离子通道抑制剂)阻断[18]。后续的工作进一步证明了重组脂质体下的Piezo有机械敏感性[19]。
至此,Piezo蛋白可以被名正言顺的称为“机械门控Piezo离子通道”。
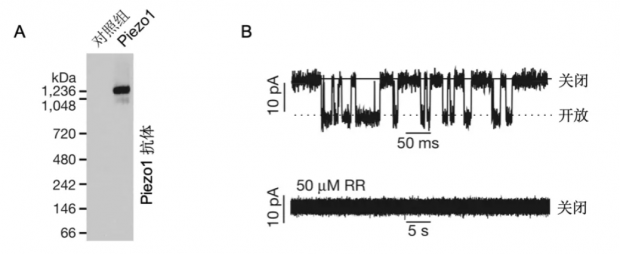
图5. Piezo1是离子通道
A,纯化的Piezo1蛋白的免疫印迹电泳图(western blot),图中黑色条带即为Piezo1蛋白的条带。B,上图,将纯化后的Piezo1蛋白重组到人工脂质体上后可记录到通道开放事件。下图,该通道开放事件可被广谱阳离子通道阻断剂钌红(RR)所阻断。(改编自Coste and Xiao, et al., 2012, Nature[18])
眼见为实——电子显微镜下的三叶螺旋桨
Piezo通道在进化上是保守的,从植物到动物都有这一蛋白[1]。但在同一个体内,它和其他的蛋白都不相似。那这个全新的蛋白长什么样呢?它究竟有怎样的架构,竟能感知机械力?以及,它是如何传递机械力,来打开通道的?
单单基于氨基酸一级序列的诸多猜测和实验都不如亲眼所见。此时,Piezo研究呼唤结构生物学登上舞台。
在首篇Piezo论文(2010年)问世后的两三年,冷冻电子显微镜技术发生了质的飞跃。贝叶斯方法的引入和直接电子检测相机(DDD)的使用,让冷冻电镜能够解析近原子分辨率的蛋白质三维结构。2013年,加州大学旧金山分校(UCSF)的程亦凡博士率先使用冷冻电镜解析了TRPV1离子通道(2021年诺贝尔生理或医学奖的另一主角)近原子分辨率结构[20],这为其他通道结构的解析打入了一针强心剂。
2013年,肖百龙博士来到清华大学组建实验室,恰逢施一公、王宏伟等人布局、搭建了强大的冷冻电镜平台。肖百龙决定与清华的结构研究者们合作解析Piezo通道的三维结构。并率先获得了Piezo1的中等分辨率结构[21]。冷冻电镜技术的突破同样吸引了Ardem和Rod Mackinnon(2003年诺奖得主)的目光,二人随即也投入到Piezo结构解析的工作中。一切都如此顺理成章——合适的人,合适的时间,合适的技术手段——接下来的几年,三方(肖百龙/李雪明,Ardem Patapoutain/Andrew B. Ward,Rod Mackinnon)几乎同时获得了Piezo1的高分辨率结构[22-24],而Piezo2的高分辨率结构也随后为肖/李合作解出[25]。
Piezo的造型十分独特。它是由三个单体组成的三叶螺旋桨状结构,多达114次跨膜,是已知跨膜次数最多的一类大型膜蛋白。蛋白单体前36次跨膜以每4个跨膜区组成一个单元,共有9个重复性的单元,最后两个跨膜区被认为形成孔道。蛋白整体由外周桨叶部分向中心凹陷成穹顶状(直径28nm,深10nm),中心孔道区域上方有一胞外的帽子结构域(Cap),细胞内侧有一9nm长的长杆结构(Beam),将桨叶外周连接到中心孔道的胞内部分[21-25]。
著名物理学家费曼说过:“你只要看到它,你就能回答许多基本的生物学问题。”
Piezo通道这般别致的结构能告诉我们什么呢?
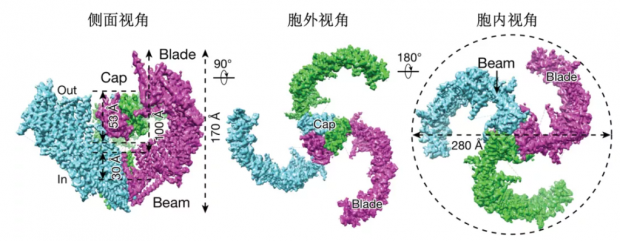
图6. Piezo2冷冻电镜三维结构卡通图(改编自Wang, et al., 2019, Nature[23])
逐鹿中原——Piezo通道如何感知力?
结构决定功能,世间万物大抵如此。
Piezo这个三叶螺旋桨打开过程中发生了怎样的变化?研究人员有理有据的“脑洞大开”。
Piezo的外周桨叶能将周围的细胞膜向内扭曲凹陷,而其他膜蛋白周围的膜成分一般是平的,这一异乎寻常的现象让研究人员们猜测:机械力作用在Piezo上的时候是不是会将该凹陷展平呢[26]?2003年诺奖得主Rod Mackinnon的课题组发表了两篇文章,论证了展平的可行性以及模型[24, 27],之后还利用原子力显微镜进一步验证[28]。
依托结构导向的功能机制研究,肖百龙课题组发表了系列文章来揭示Piezo通道的机械门控机制,并提出了系列模型假说,譬如“杠杆模型”假说:胞内的长杆结构可作为杠杆撬开孔道[22, 29]。此外,蛋白上各个结构域的潜在作用也被逐步揭示:桨叶可以作为机械力传导模块[30];锚定区(anchor)后的连接(linker)作用很关键:内质网蛋白SERCA可通过它抑制Piezo[31],而E-cadherin蛋白可通过它将Piezo和细胞骨架联系在一起而增强Piezo功能[32];帽子区可感知、传导机械力,并且决定了Piezo通道从打开到失活过程的快慢[25, 30];长杆区之后的“门塞”序列可通过“门塞和闩锁”机制完成Piezo三个胞内侧向通路的协调门控[33]。
期待有朝一日,我们能获得Piezo通道从关闭到开放再到失活等全部状态下的结构以及在细胞膜上的原位结构,能将上述各种学说囊括入一个“大统一”框架内。
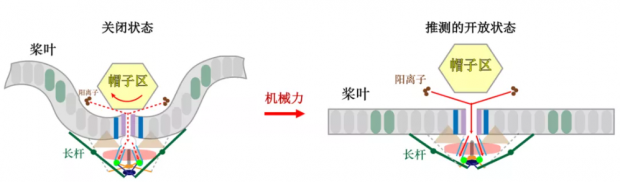
图7. 推测的Piezo通道响应机械力而开放的模型
Piezo通道的侧面视角,为便于查看,Piezo三聚体简化为两个亚基。红色虚线箭头和实线箭头分别代表离子不通透和通透的状态。(改编自Jiang, et al., 2021, Trends Biochem Sci[34])
(未完待续)
参考文献
1. Coste, B. et al. Piezo1 and Piezo2 are essential components of distinct mechanically activated cation channels. Science 330, 55-60 (2010).
2. Hargrave, P.A. et al. The structure of bovine rhodopsin. Biophys Struct Mech 9, 235-244 (1983).
3. Nathans, J. & Hogness, D.S. Isolation, sequence analysis, and intron-exon arrangement of the gene encoding bovine rhodopsin. Cell 34, 807-814 (1983).
4. Buck, L. & Axel, R. A novel multigene family may encode odorant receptors: a molecular basis for odor recognition. Cell 65, 175-187 (1991).
5. Hoon, M.A. et al. Putative mammalian taste receptors: a class of taste-specific GPCRs with distinct topographic selectivity. Cell 96, 541-551 (1999).
6. Ranade, S.S., Syeda, R. & Patapoutian, A. Mechanically Activated Ion Channels. Neuron 87, 1162-1179 (2015).
7. Sukharev, S.I., Blount, P., Martinac, B., Blattner, F.R. & Kung, C. A large-conductance mechanosensitive channel in E. coli encoded by mscL alone. Nature 368, 265-268 (1994).
8. Corey, D.P. & Hudspeth, A.J. Response latency of vertebrate hair cells. Biophys J 26, 499-506 (1979).
9. Martinac, B. & Poole, K. Mechanically activated ion channels. Int J Biochem Cell Biol 97, 104-107 (2018).
10. Guharay, F. & Sachs, F. Stretch-activated single ion channel currents in tissue-cultured embryonic chick skeletal muscle. The Journal of physiology 352, 685-701 (1984).
11. Hoffman, B.D., Grashoff, C. & Schwartz, M.A. Dynamic molecular processes mediate cellular mechanotransduction. Nature 475, 316-323 (2011).
12. Christensen, A.P. & Corey, D.P. TRP channels in mechanosensation: direct or indirect activation? Nat Rev Neurosci 8, 510-521 (2007).
13. Arnadottir, J. & Chalfie, M. Eukaryotic mechanosensitive channels. Annu Rev Biophys 39, 111-137 (2010).
14. Chalfie, M. & Au, M. Genetic control of differentiation of the Caenorhabditis elegans touch receptor neurons. Science 243, 1027-1033 (1989).
15. Huang, M. & Chalfie, M. Gene interactions affecting mechanosensory transduction in Caenorhabditis elegans. Nature 367, 467-470 (1994).
16. Walker, R.G., Willingham, A.T. & Zuker, C.S. A Drosophila mechanosensory transduction channel. Science 287, 2229-2234 (2000).
17. Patel, A.J. et al. A mammalian two pore domain mechano-gated S-like K+ channel. EMBO J 17, 4283-4290 (1998).
18. Coste, B. et al. Piezo proteins are pore-forming subunits of mechanically activated channels. Nature 483, 176-181 (2012).
19. Syeda, R. et al. Piezo1 Channels Are Inherently Mechanosensitive. Cell reports 17, 1739-1746 (2016).
20. Cao, E., Liao, M., Cheng, Y. & Julius, D. TRPV1 structures in distinct conformations reveal activation mechanisms. Nature 504, 113-118 (2013).
21. Ge, J. et al. Architecture of the mammalian mechanosensitive Piezo1 channel. Nature 527, 64-69 (2015).
22. Guo, Y.R. & MacKinnon, R. Structure-based membrane dome mechanism for Piezo mechanosensitivity. eLife 6 (2017).
23. Saotome, K. et al. Structure of the mechanically activated ion channel Piezo1. Nature 554, 481-486 (2018).
24. Zhao, Q. et al. Structure and mechanogating mechanism of the Piezo1 channel. Nature 554, 487-492 (2018).
25. Wang, L. et al. Structure and mechanogating of the mammalian tactile channel PIEZO2. Nature 573, 225-229 (2019).
26. Murthy, S.E., Dubin, A.E. & Patapoutian, A. Piezos thrive under pressure: mechanically activated ion channels in health and disease. Nature reviews. Molecular cell biology 18, 771-783 (2017).
27. Haselwandter, C.A. & MacKinnon, R. Piezo's membrane footprint and its contribution to mechanosensitivity. eLife 7 (2018).
28. Lin, Y.C. et al. Force-induced conformational changes in PIEZO1. Nature 573, 230-234 (2019).
29. Wang, Y. et al. A lever-like transduction pathway for long-distance chemical- and mechano-gating of the mechanosensitive Piezo1 channel. Nature communications 9, 1300 (2018).
30. Zhao, Q. et al. Ion Permeation and Mechanotransduction Mechanisms of Mechanosensitive Piezo Channels. Neuron 89, 1248-1263 (2016).
31. Zhang, T., Chi, S., Jiang, F., Zhao, Q. & Xiao, B. A protein interaction mechanism for suppressing the mechanosensitive Piezo channels. Nature communications 8, 1797 (2017).
32. Wang, J., Jiang, J., Yang, X., Wang, L. & Xiao, B. Tethering Piezo channels to the actin cytoskeleton for mechanogating via the E-cadherin-β-catenin mechanotransduction complex. bioRxiv (2020).
33. Geng, J. et al. A Plug-and-Latch Mechanism for Gating the Mechanosensitive Piezo Channel. Neuron 106, 438-451 e436 (2020).
34. Jiang, Y., Yang, X., Jiang, J. & Xiao, B. Structural Designs and Mechanogating Mechanisms of the Mechanosensitive Piezo Channels. Trends Biochem Sci 46, 472-488 (2021).
作者介绍

刘文豪,清华大学生命学院2016级PTN项目博士生,师从肖百龙教授。博士期间主要从事机械门控Piezo通道结构功能关系的研究,曾以共同第一作者在Nature、Neuron发表文章揭示Piezo通道机械门控过程的“双门控”机制和“门塞和闩锁”机制。
话题:

0
推荐
财新博客版权声明:财新博客所发布文章及图片之版权属博主本人及/或相关权利人所有,未经博主及/或相关权利人单独授权,任何网站、平面媒体不得予以转载。财新网对相关媒体的网站信息内容转载授权并不包括财新博客的文章及图片。博客文章均为作者个人观点,不代表财新网的立场和观点。
 京公网安备 11010502034662号
京公网安备 11010502034662号 {kind=link}