格兰德·艾伦(Garland E. Allen)是生物学领域思想史研究的重要人物之一,于2017年荣膺科学史领域最重要的奖项——萨顿奖。艾伦教授对遗传演化有深刻理解,本文根据他2013年发表的一篇科学史论文进行了节选编译,文章通过考察现代遗传学之父、美国生物学家托马斯·亨特·摩尔根(Thomas Hunt Morgan)由胚胎学家转变为遗传学家的案例,佐证了科学研究的特质——“错误”的观念也可以很有用处,精彩还原了遗传学发展史上激动人心的一个片段。

Garland E. Allen
编译|李峰
在教科书中或课堂上,人们习惯于以当下的标准来解释过去那些“错误观点”,指出谬误所在,却对这些谬误产生的背景乃至受到追捧的原因避而不谈。然而,恰恰是那些后来被证伪的假说,隐含了科学研究中的诸多精彩细节和内幕,也真实呈现出科学始终是“在争议中前行”的动态发展的本质。
20世纪前十年,后来被誉为“现代遗传学之父”的美国生物学家托马斯·亨特·摩尔根(Thomas Hunt Morgan)强烈反对当时最重要的三种观点:达尔文自然选择理论、遗传的染色体理论以及孟德尔遗传学。但他随后改变了对这三种理论的态度,发展出充满生命力的研究体系,铺平了统一遗传学和达尔文演化理论的道路,这就是30年代的“综合进化论”(the evolutionary synthesis)。1933年的诺奖得主、被誉为“现代遗传学之父”的美国进化生物学家托马斯·摩尔根(Thomas Hunt Morgan,1866.9.25-1945.12.4)是个难以捉摸的人物。英国遗传学家威廉·贝特森(William Bateson)称之为“完全没有一点自负”的人——特别是碰到他不懂的事的时候——并且美籍俄裔遗传学家西奥多修斯·杜布赞斯基(Theodosius Dobzhansky)认为他是怀疑论者、特立独行的人,经常语出惊人,而又始终保持绅士风度。这是摩尔根众多个性特点中的两例,但是非常适合考察他在遗传学研究中的角色。

摩尔根
摩尔根的生物学训练开始于约翰霍普金斯大学,学习形态学。那时候形态学的核心是运用胚胎发育中的特征来构建演化上的关联(系统发育)。形态学概念充满想象又有意思,可以产生出各种不同的解释,但是没有一种能够被严格地检验。像很多同时代人物一样,摩尔根彻底厌倦了本质上是极度臆测的形态学研究工作。特别是在1890年代,到意大利拿波里动物研究所(Naples Zoological Station)访学之后,他熟悉了胚胎生物学中新兴的实验研究工作,很快成为实验生物学的热心支持者。在实验中可以人为干涉,每次控制一个变量,从而严格地检验假说。
质疑主流理论题
1 反对达尔文理论
1903年,摩尔根在他的《演化与适应》一书中阐明了对达尔文自然选择理论的反对。需要指出的是,摩尔根完全赞同演化过程——也就是逐代改良(descent with modification)。因为在他看来,自然选择的机制,作用于小的、独立的变异,永远不能产生新种。虽然达尔文依赖于把人工选择类比于自然选择,以此强调种的可变性,但现在我们都知道了,经过育种者几百年来的选育,从没产生过任何“新种”。作为一种替代方案,二十世纪初,摩尔根曾经特别着迷荷兰植物学家德佛里斯的突变理论。德佛里斯认为一代的大幅度改变就可以产生新种,我们今天称之为“大突变”(macromutations)。摩尔根对德佛里斯的主张印象深刻,因为他对温室中的模式生物月见草 (Oenothera)突变体进行了研究,突变体可以通过实验的方法来检验。
摩尔根认为达尔文观点中还有其他问题。比如任何性状要想有适应性,它必须是完全形成的。如果是不完全形成的器官,比如脊椎动物的眼,就会适应性很低,或者完全没有适应性。因此,在这些很多小的步骤中怎么可能实现演化呢,没有哪个微小的突变对个体来说有任何适应性价值。
2 反对染色体遗传说
就像厌恶臆测的方法而反对自然选择一样,摩尔根也反对当时的染色体遗传理论。当时大多数生物学家意识到,有丝分裂和减数分裂中复杂而有规则的染色体运动预示着某些重要的东西,但是这些东西到底是什么则完全不清楚。最流行的观点是这一定跟遗传有些关联,但是直到1909年摩尔根对这种说法仍然十分怀疑。比如,他指出在两次细胞分裂的间期,棒状染色体消失在视野中。摩尔根问道,如果它们在每次细胞周期的末尾都解体,染色体又如何能维持细胞代际间的遗传稳定?我们怎么知道下一次循环中,染色体构建出了同样的结构呢?
对于摩尔根来说,更加矛盾的地方是当时的另外一个重要案例——性别决定问题。1904到1905年,哥伦比亚大学生物系主任艾德蒙德·威尔森和布林马尔女子学院的奈特·史蒂文斯确定了性别的决定是由一对染色体的分布控制的,就是所说的X染色体或者副染色体。但摩尔根觉得这种解释自相矛盾:绝大多数动物中雄性动物有一条X染色体,而雌性个体有两条;但是鸟类、蝴蝶和蛾子三类动物中情形相反。同一种染色体分布如何能够在一类情形时决定雄性,而在另一类又决定雌性?
3 反对孟德尔遗传
在他批评染色体学说的同一年,他还写文章猛烈批判快速发展的孟德尔遗传理论(Morgan 1909)。他发现孟德尔学派像其他老一派形态学家一样,随意地毫无节制地臆测。例如,为了解释预期的孟德尔比例中出现的异常(比如上位作用),孟德尔主义者简单引入不同的因子:“如果一个因子不能解释现象,那就引入两个因子;如果两个还不够用,三个也许就管用了……”,摩尔根称之为“高级杂耍”。而且,摩尔根还注意到,决定性状的孟德尔主义“因子”散发着就形态学预成论的气息,把发育过程(包括分化)归结为假定的“完全决定的”微小个体,或者来自精子,或者来自卵子,以此回避了成体中的性状到底是如何形成的问题[1]。显然摩尔根的这一质疑是对的。孟德尔最初的理论完全没有提到胚胎发育,只是一个描述一代传给另一代的杂交过程中性状分布的理论。
抛开这些反对自然选择、染色体、孟德尔遗传理论的具体理由,是什么让摩尔根在很短时间内对三种理论改变立场?
果蝇带来的改变
01 摩尔根的第一次观念改变
1908年,摩尔根开始在实验室培养果蝇,想看看是否能在动物中发现德佛里斯式的新种水平大突变。摩尔根让一个学生在他的实验动物学课程中使用果蝇,暗中连续培养69代,来检验获得性性状可遗传的“新拉马克主义”实验结果是否定的。然而在这一过程中,摩尔根确定果蝇是个很好的实验体系,它易于获取,培养廉价,不占空间,每2-3周繁殖一代。但是然而直到1909年冬天,他没有得到任何重要结果。在1910年1月,摩尔根发现了几个突变体,最显眼的突变体是“三叉戟” (Carlson 2004: pp. 170–171)。与野生型杂交,这一性状看起来像是隐性的,但是在第二代,有很多变种,很难建立起纯系。1910年春天,又出现两个突变体:Olive(2月)和Speck(3月)。但是这两个突变体看起来都不是明显的孟德尔定律预期的3:1或者9:3:3:1比例,这也印证了摩尔根早先的观点,孟德尔的不连续的显隐分类是很武断的。在这一时期摩尔根也开始重新考虑他对于染色体理论的观点。因为威尔森和史蒂文斯的结果清楚地表明性别与染色体构型有着某种关联。即使有摩尔根质疑的那些问题,这种关联太有规律了,不大可能仅仅是人为的假想。另外,1905-1908 年,植物学家和动物学家的细胞学研究清楚地说明雄配子(花粉或精子)有两种类型,差别在于是携带了X染色体,还是携带了怪异的对应的Y染色体。总之,对于大多数生物来说,看起来都是雄配子决定性别。这就能轻松解释绝大多数生物中观察到的雌雄比保持1:1。性别决定研究与西奥多·鲍维里新进发表的“染色体特性”也很符合,也就是每一对染色体可能决定一对性状或者发育过程 (Boveri 1907)[2]。
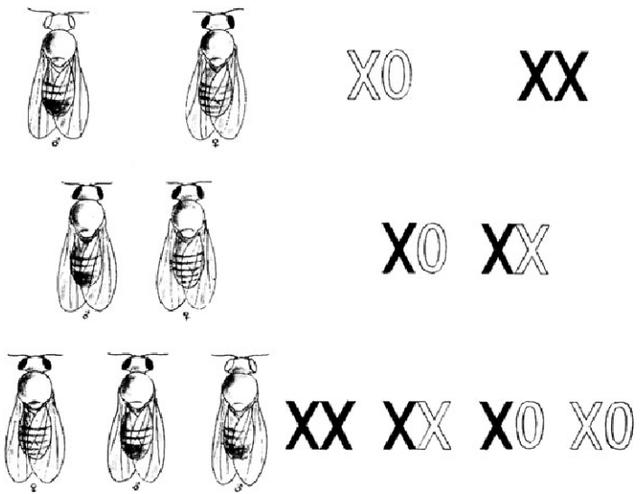
图1 孟德尔白眼雄果蝇和红眼雌果蝇杂交实验的早期呈现方式(1913年)。X染色体或者副染色体用实线(红眼)和白色(白眼)表示。这一时期,还不清楚雄果蝇有Y染色体,所以雄性染色体组成用XO表示。(Morgan 1913: p. 19)。
但是在1910年5月,摩尔根在他的培养瓶中发现了一只白眼雄果蝇,于是把它与一只正常红眼雌果蝇杂交。后代全部是红眼,结果显示红色相对于白色是显性。后代雌雄比1:1,像预期的一样。但是当他把F1代的雌雄果蝇杂交,F2代产生了3:1的红白比,看起来是符合了孟德尔遗传比例,但奇怪的是,隐性性状只出现在雄性果蝇 (图1)。实际颜色比例非常符合孟德尔遗传比例这一事实,让摩尔根接受了孟德尔方案,至少是初步接受了。与此同时,E.B. 威尔森明确支持把染色体与孟德尔因子联系起来。在6月,他给Science杂志投了如今非常著名的那篇文章“果蝇中的限性遗传”(Sex-limited Inheritance in Drosophila)。这篇文章有两个有意思的特点,说明摩尔根对于是否把孟德尔因子和染色体直接联系起来,仍然犹豫不决。首先是题目:“限性”(sex-limited)表明了白眼主要与雄性相关,或者说限定于雄性,但是机制尚不明确。其次,此时摩尔根描述亲代、子代基因型的方式,暴露出他仍然不愿意把决定眼色的因子在物理上定位于X染色体上。他用现在已经过时的标记符号表示眼色因子(W代表白色,R代表红色),用X染色体代表性别(XX代表雌性,X代表雄性)[3],把眼色和性别当做不同的性状来独立分类[3]。他用下面的方式表示F1和F2代的杂交结果:

F1代个体间进行杂交

这里红眼雌果蝇有两种类型:一种是纯合体(RR),另一种是杂合体(RW);但是对于雄果蝇有两种基因型和两种表型:红眼雄果蝇是杂合体(RW),白眼雄果蝇是纯合体(WW)。现在摩尔根左右为难,他完全知道“为了获得这样的结果,必须假设……当F1代雄性红眼果蝇(RWX)形成两类精子的时候,R和X在一起,否者结果就不符合(如图所示)”。在最后,摩尔根假设R和X总偶联在一起,把眼色因子和X染色体看作是独立的实体。也许他已经考虑用其他方法描述杂交结果,但此时他还沿用以上符号,并把眼色和性别看作独立的因子。
承认R和X偶联在一起给摩尔根制造了问题:一方面是为什么F1代红眼雄蝇中这两个因子永远一起分离?显然摩尔根对这个假设感到不舒服,因为这明显是临时的:这是用来解释为什么他在F2代中没有找到白眼雌果蝇的。在1910年,威尔森甚至建议摩尔根,最简单的解释就是假设眼色因子与X染色体在物理上相联系。但是摩尔根觉得这种观点还是太过臆测。他开始改变想法了,但是此时他还不能彻底赞同。
那个夏天,培养中出现了更多的突变体,出现了残翅、袖珍翅和黄身这些像白眼一样是“性别限定”的性状。新发现的很多限性遗传性状,让威尔森建议的简明解释——这些神秘的性状,独立地与X染色体一起分离——变得很难想象。另一方面,如果它们确是X染色体上的一部分,就顺理成章地解释了为何与性别相关联。因此摩尔根心中产生了染色体上具有多个孟德尔因子的想法。所以,1915年摩尔根发展了伴性遗传的想法,而且扩展到了更广泛的联系,让他用新的方式标示白眼果蝇杂交(图2)。
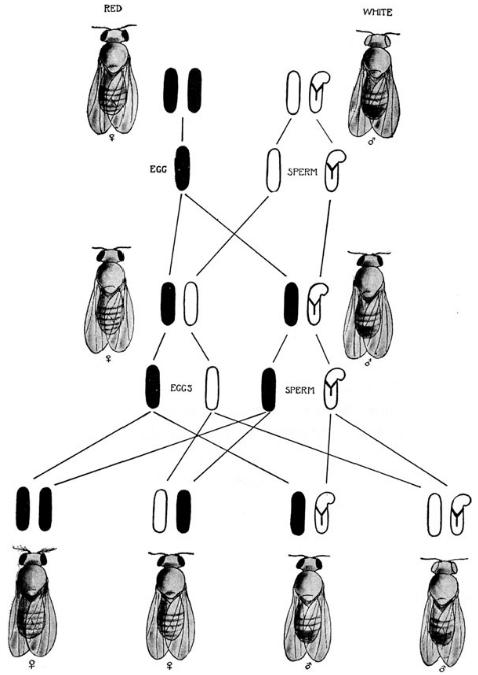
图2 在这个示意图中,不同于之前的版本,颜色因子直接与X染色体实体相联系。黑色染色体携带红眼因子,白色染色体携带白眼因子。Y染色体此时已经被证实,用Y来标识,上端弯曲 (Morgan et al. 1915: p. 17) 。
02 更多数据:发现连锁与互换
此时的摩尔根接受了孟德尔遗传和染色体理论,以及二者实体上相互关联的判断。但他面临着另一个早就广为人知的动植物育种问题,包括达尔文本人也困惑不解:为什么相互连锁的两个性状,偶尔也会被分开,或者说打断,后代中两个性状就不会再一起出现,但是可能会重新组合(AB和ab的组合偶尔会产生出Ab和aB)?
1906年,英国的威廉·贝特森发表了有关这一问题的报告,他用香豌豆为材料进行实验 (Bateson 1906)。花的颜色蓝色(B)相对红色(b)是显性,花粉外型长(L)相对于圆(l)是显性,而花的颜色与花粉外型表现出连锁关系,所以BL纯合体与bl纯合体杂交后 ,F1代表型应该全部是蓝花、长花粉,但是它们应该都是杂种或者说杂合体(BbLl)。F1代的杂合体自交,F2代大多数(87.4 %)是连锁的,表型像它们的祖父辈一样,也就是蓝长和红圆,但是也有小部分(12.6 %)是重组的,也就是蓝圆和红长。像摩尔根一样,贝特森也怀疑染色体学说,他提出了更复杂的“吸引与排斥”(attraction and repulsion)理论来解释这种现象。摩尔根认为贝特森的想法都很复杂,而且没有任何实际支持。
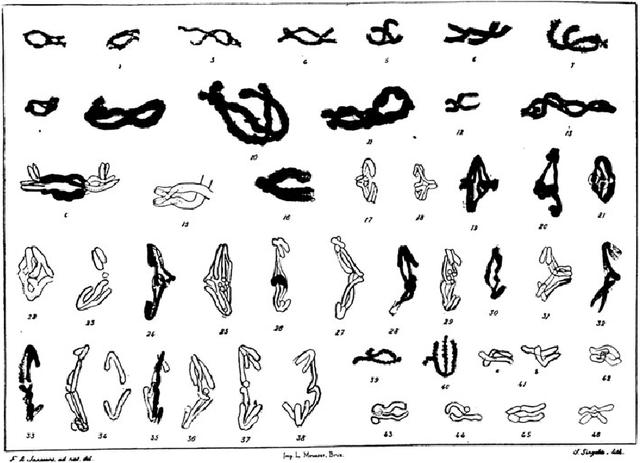
图3 1909年比利时细胞学家詹尼斯在论文中描述的,不同阶段的染色体交叉(准确的说应该是同源染色体对复制后,四分体状态的姐妹染色单体)。詹尼斯认为在姐妹染色单体交叉的地方,可能发生同源染色体之间的断裂和重连,产生包含着父母双方的互补片段的姐妹染色单体。(仿自詹尼斯,摩尔根发表于1913: p. 93)
就在这个时候,威尔森又一次为故事提供了关键情节:他让摩尔根关注比利时细胞学家佛朗斯·阿方斯·詹尼斯在1910年发表的文章。文中数据展示了减数分裂前期纠缠在一起的染色体(图3)。对于摩尔根来说,詹尼斯的图暗示了,在缠绕之前,同源染色体之间(已经完成复制的姐妹染色单体)会发生断裂,同源的末端会重连,这为同一染色体上的因子之间解偶联和重组提供了解释(图4)。他很快意识到,两个孟德尔因子在染色体上是分离的,在两者之间经常会发生断裂,因此会在后代中表现出很大比例的重组表型。之前20年间积累的对染色体以及染色体运动的实际观察,为解释遗传机制提供了基础。这种机制不需要发明离奇的吸引和排斥:连锁和互换都可以用同样的机制解释,而且与已知的染色体结构和功能相符合。细胞学和育种这两条线的证据汇集在一起,造就了1915年著名的“孟德尔-染色体遗传理论”。
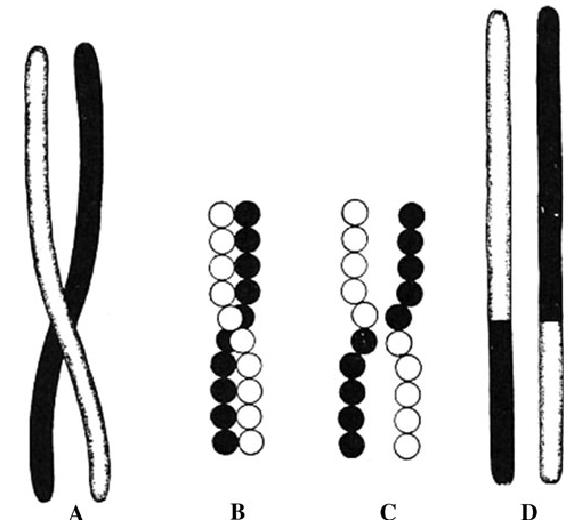
图4 表示同源染色体(更确切的说是姐妹染色单体)联会的示意图,摩尔根1915, a和 d姐妹染色单体交换了同源部分; b 和 c表示了由单个基因组成的染色体。黑白两色分别代表同源染色体来自亲本中的父方母方 (Morgan et al. 1915: p. 60) 。
在这一时期,摩尔根意识到他一个人跟不上快速增长的果蝇研究工作了,于是开始培养有天赋的本科生。在他实验室日后毕业的学生中,有三人最为著名:阿尔弗雷德·斯特蒂文特、卡尔文·布里奇和赫尔曼·穆勒。摩尔根越来越依赖这些学生从事日常工作,去揭示业已结合在一起的孟德尔-染色体范式之间的内在联系。举个例子,1911年,摩尔根与斯特蒂文特讨论重组频率与染色体上两个基因间距离的问题时,遗传图谱的想法诞生了。斯特蒂文特回忆,当晚他回家后,彻底被这个想法迷住了,忘记了写家庭作业,并绘制了第一幅染色体图谱:染色体上连锁的6个基因(Sturtevant 1965: 47)。之后染色体图谱绘制工作迅速展开,最终在果蝇四对染色体上定位了数百个基因。
“孟德尔-染色体”范式的建立
摩尔根的学生组成了充满热情的团队,推动染色体图谱工作,并开始详细阐述一整套遗传法则,拓展和澄清了孟德尔主义模型,延伸到了更多的问题,比如基因-基因相互作用(上位效应,epistasis)、基因多效性(pleiotropy)、染色体突变(插入、移位)、性别决定的染色体机制(布里奇)、同一位点的复等位基因(斯特蒂文特),以及最终的人工诱变(穆勒)。在20世纪前30年,孟德尔-染色体理论,以及与其相联系的染色体图谱技术带来的众多问题,把遗传学推到了生物学最重要的前沿领域。

图5 1919年夏天,在马萨诸塞的伍兹霍尔海洋生物学实验室,摩尔根和几个实验室成员讨论实验结果。|供图:伊莎贝尔·摩尔根·蒙坦
果蝇研究还是现代生物学中实验室团队研究的代表。从社会学角度来看,这是一段很有意思的历史。在1911-1915年间,团队像一个整体,工作气氛和关系是很开放和自由的,日后的内部争斗还未露端倪。大家很享受夏天在MBL开的实验室聚会和非正式“实验室会议” (图5)。实际上,他们第一次推动了新的科学研究形态—团队研究。在19世纪和20世纪早期,生物中的科学研究工作主要由个人进行研究。论文和专著都由一个研究者撰写,多个共同作者的情况比较少见。绝大多数欧洲实验室受正统规则主导,等级分明。摩尔根的研究组则打破了常规,以更加自由流动的方式运作,老师参与少,由学生做主。
1915年,研究团队的成果首次完整地总结在《孟德尔遗传机制》一书中,作者是摩尔根、斯特蒂文特、布里奇和穆勒。这本书现早已经成为经典,写作精炼,插图精美,被翻译为多种语言,使得遗传学研究中的新成果引起了全世界的关注。
随着绘制遗传图谱的工作的深入,孟德尔因子与染色体之间的关系越来越清晰。在果蝇中出现了四个连锁群,与染色体对数一致,说得更明白一些,就是每对染色体的实际大小与遗传因子数量大致相同 (图6)。事实上,这种关系变成了最强的一条证据,让任何怀疑者都相信基因就是染色体上的实体部分。在上世纪20年代末期和30年代,康奈尔大学的罗林斯·埃莫森在玉米中,哈佛大学的凯斯特利在老鼠中都建立了连锁群。20世纪上半叶,孟德尔-染色体理论已经成为了经典遗传学的基础。
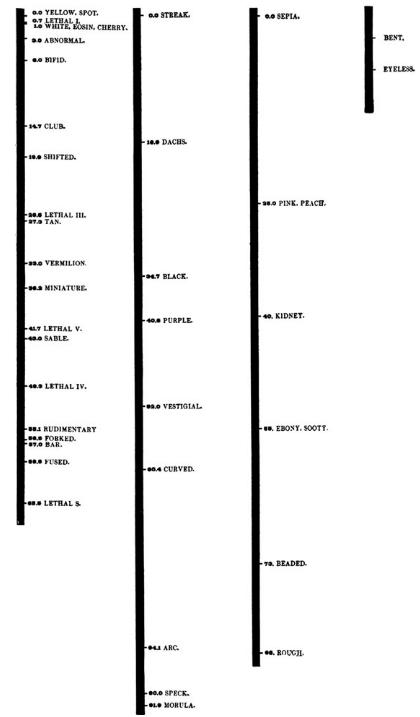
图6 早期(1915)果蝇四个连锁群的染色体图谱示意图。染色体从左到右依次为I (X染色体), II, III and IV号染色体。连锁群的数量由杂交结果确定,与细胞学观察到的染色体对数目一致, (来自摩尔根 Morgan et al. 1915: 封面插图) 。
大约在《孟德尔遗传机制》一书出版的时候,摩尔根第三次改变了立场,开始接受这种作用于小的、不连续的孟德尔变异的自然选择,当作重新流行起来的达尔文演化观念的基础。在他的《进化论评论》一书中,摩尔根指出不连续的变异,以及由不同基因相互作用产生的一系列梯度变异(称之为数量遗传),是自然选择发生作用的微小个体差异的来源 (Morgan 1916: pp. 85–87)。很多年后,据穆勒回忆,他和实验室的其他人同摩尔根争论过演化问题,最后摩尔根认同孟德尔式变异可以为演化提供原始材料 (Allen 1968: pp. 135–137)。这一认识,移走了摩尔根对于自然选择看法中最后一块绊脚石。虽然没有像后来建构综合进化论的罗纳德·费舍尔或者休厄尔·赖特那样提出数学公式,但是在选择作用于微小的不连续遗传突变上,摩尔根的观点与他们一致,认为在传给下一代时突变的数量可以增加或减少。
对教育的启示
在这篇文章中,我着重强调了1900-1916年之间,摩尔根早期研究中的几个特点,这些特点展示了:(1)在任何科学发展过程中,那些被(同时代或者后来者)认为是走了弯路或者错误的观点的重要性,以及向学生讲授这些观点的价值;(2) 科学是个社会过程,发生在特定时间、地点和特定的社会-经济背景下。科学家们在研究工作中,除了单纯的“发现真理”,还有着各种的现实目的。
有关第一点,我们已经看到,在概念上,摩尔根对于自然选择、染色体遗传理论和孟德尔的遗传传递模型上一开始都做出了错误的评价,但是他都有充分的理由。对于摩尔根来说,孟德尔理论和染色体学说不仅是反映了早期的、被抛弃的胚胎学预成论的回光返照,而且回避了最重要的问题。对于摩尔根这样的胚胎学家,胚胎发育过程中这些性状是如何产生的才是最重要的问题。这个问题仍然是今天的生物学最前沿的问题,构成了新兴的演化发育生物学(evo-devo)领域的核心。
关于第二点,20世纪头十年中,摩尔根是年轻生物学家中的领袖,他想把生物学带出形态学臆测的困境,构建在像物理化学一样坚实的基础上。摩尔根和其他科学家想要在仍被物理学统治的领域中争取到合理的一席之地的努力,是生物学发展历史上至关重要的部分。
以上这些该如何帮到今天实际的生物学教学中呢?最简单的一级是,好的科学家会承认错误,改变自己的观点,实际上,这也是科学作为人类活动的一种力量体现。更高一级地说,这些案例可以促进对本文讨论的三种理论的理解,通过列举各种摩尔根的反对意见,让学生用对这些问题的现有理解,做出自己的回答。这种方法必然会引起很多讨论。这会强化这样的观点——知其然,也知其所以然。直面这些理论的反对意见,可以帮助学生更加深刻地理解那些如今我们自认为了解的东西。
在关于摩尔根的案例中,我试图展示出科学像流水的一面,科学没有固定的方法或者路径。如果能为学生展现一个动态的过程,错误的理论也会像正确的一样有用。这样在我们为下一代人展现科学时,会更加令人激动,更加娴熟自如。
注释
[1] 虽然1900年以后,差不多所有胚胎学家都已经拒绝了18世纪早期的预成论,但是它仍然像飘荡在野地的鬼魂,一旦有机会,随时准备借尸还魂。
[2] 鲍维里用实验确定个体特征的想法非常天才:在海胆卵的培养皿中加入高浓度精子,可以出现双受精的卵。在这些双受精个体中有丝分裂的染色体分布是不正常的,挑选出后,继续发育,会产生不同种类的畸形,畸形的种类取决于哪条染色体缺失。特定的胚胎生理畸形和缺失特定染色体之间的有规律联系,让鲍维里相信,用他自己的话来说,“正常发育所需的不是确定的染色体数目,而是染色体的特定组合,这无异于说明了每个单独的染色体一定具有不同的特性。”(鲍维里1902,沃勒尔1968年翻译重印)
[3] 那时仍不知道果蝇雄性个体有Y染色体;在很多种类的昆虫中,雄性是XO,而雌性是XX,所以认为果蝇也是这样(参见图1,摩尔根在1913年的早期性别限定的标识方式)。到了1915年,已经清楚雄性果蝇确实有Y染色体,应该标识为XY。
本文编译自 Garland E. Allen, How Many Times Can You Be Wrong and Still Be Right?
T. H. Morgan, Evolution, Chromosomes and the Origins of Modern Genetics,Sci & Educ.
DOI 10.1007/s11191-013-9664-8
特 别 提 示
1. 进入『返朴』微信公众号底部菜单“精品专栏“,可查阅不同主题系列科普文章。
2. 『返朴』提供按月检索文章功能。关注公众号,回复四位数组成的年份+月份,如“1903”,可获取2019年3月的文章索引,以此类推。
版权说明:欢迎个人转发,任何形式的媒体或机构未经授权,不得转载和摘编。转载授权请在「返朴」微信公众号内联系后台。
特 别 提 示
1. 进入『返朴』微信公众号底部菜单“精品专栏“,可查阅不同主题系列科普文章。
2. 『返朴』提供按月检索文章功能。关注公众号,回复四位数组成的年份+月份,如“1903”,可获取2019年3月的文章索引,以此类推。
欢迎个人转发,任何形式的媒体或机构未经授权,不得转载和摘编。转载授权请在「返朴」微信公众号内联系后台。

0
推荐
 京公网安备 11010502034662号
京公网安备 11010502034662号 {kind=link}