阅读:0
听报道
许多做出了伟大发现的科学家在回忆当年的情况时,都会说:“我偶然发现……”也许是一个反常的现象,也许是一串异常的数据,又或者是一种从没见过的奇怪事物。接着,根据教科书或史书上的记载,他们就顺理成章地做出了伟大发现,仿佛真理只是散落在草丛里闪闪发亮的宝石,运气好的话弯腰捡起就能发大财。
当然,事实并非如此。实验中出现的各类异常,很容易被当作杂音忽略,当作错误筛除,当作极端值排除。需要勇气和眼光,才敢去、才能去冒险挖掘异常背后的故事。这场探险的旅程中,科学家是如何掌握线索、如果顺藤摸瓜,如何检索到数量稀少但却至关重要的文献的?他们怎样全盘思考,怎样寻找研究的切入点,怎样搞清自己走到了哪一步?今天的文章讲述了清华大学俞立老师发现和研究迁移体的历程,希望带给大家一些启发。
撰文 | 俞立(膜生物学国家重点实验室)
我现在还记得,那是2012年的一天。当时,我正在检查前一天透射电子显微镜拍出的图像。有个不寻常的东西吸引了我的眼球。那是一个位于细胞之外的囊泡,其中充满了很多更小的囊泡,看起来像一只开口的石榴。回想起来,我以前见过类似的结构,但这张图像的独特之处在于,多个囊泡集中地位于胞外,有些内部是空的,有些有少数小囊泡,还有些充满了小囊泡。这张电镜图促使我开始思考这些结构到底是什么。是死细胞的碎片?过于整洁了。是外泌体?体积太大了。是脱落小泡吗?这倒是有可能,但囊泡内部又有什么呢?很快,大家就把这种囊泡戏称为石榴体(pomegranate-like structure,PLS),以这幅电镜图像为起点,我们开始了对石榴体的研究之旅。
为了鉴定出石榴体的标记蛋白,我们首先使用亚细胞分离法分离得到石榴体,再利用质谱技术与绿色荧光蛋白(GFP)标记的方法,很快鉴定出石榴体上富集的多种蛋白。其中,四跨膜蛋白4(Tetraspanin4,Tspan4)大量富集于PLS,能够作为一个很好的标记蛋白。但真正令我们吃惊的是,Tspan4同时会标记出与一个同细胞尾端相连的膜管网络,而石榴体就位于这些膜管的分叉点或末端。它整体的外观就像一个集成电路(图1)。快速检索文献后,我们发现这些膜管是收缩丝(retraction fibers),首次发现于1963年(Taylor and Robbins, 1963)。不过,目前尚未有文献报道过PLS结构。利用绿色荧光蛋白标记的Tspan4,我们实时拍摄了石榴体的形成过程,并很快发现它与细胞迁移相关。当细胞迁移时,收缩丝被拉出,石榴体开始在收缩丝的末端或交接处生长。随着细胞的不断迁移,收缩丝断裂,石榴体从细胞上脱离。鉴于石榴体的形成依赖于细胞迁移,我们把石榴体正式命名为迁移体(migrasome)(Ma et al., 2015)。
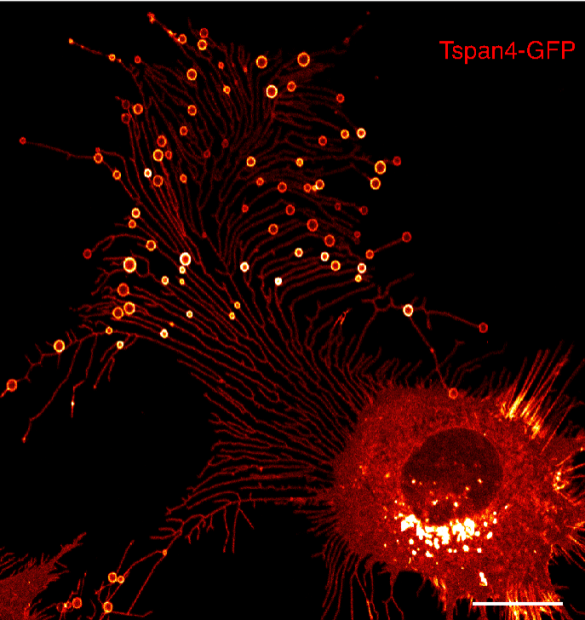
图1:L929细胞(译注:一种小鼠成纤维细胞)中的迁移体。比例尺,20微米。
在发现迁移体后,最重要的问题是迁移体有什么功能。在当时的研究阶段,我们无法真正回答这个问题,我们甚至不确定迁移体是否存在于活体内。因此我们暂时绕开这一问题,转而研究迁移体能做什么。它们是用来释放细胞质蛋白等细胞内容物的吗?的确,我们发现mCherry(译注:一种红色荧光蛋白)可以转运到迁移体中,随后通过迁移体从细胞中释放出去。我们把这一过程命名为迁移性胞吐(migracytosis)。迁移体可以介导细胞间通讯吗?我们发现一个细胞产生的迁移体可以被另一个细胞吞噬,这表明迁移体至少可能参与胞内物质在细胞之间的传递,因而可能介导细胞间通讯。虽然这些概念验证性的实验还不能告诉我们迁移体的功能究竟是什么,但这些结果让我们提出了一系列假说,以引导下一阶段的研究工作。
这些假说是:
1)迁移体产生于免疫细胞、转移的肿瘤细胞以及正在发育阶段中的细胞等迁移细胞中。
2)迁移体由确定的遗传通路调控;
3)迁移体可以介导细胞间的通讯。
在过去几年中,我们为理解迁移体所做的探索性工作主要由以上假说引导,其中一些假说目前已经得到证实。
除了假说之外,我们同样需要一个研究策略。为了回答迁移体究竟具有什么功能(这可能是最重要的问题),我们需要建立一个研究迁移体的体内模型。有了这个模型,我们就可以制备迁移体缺陷型动物,找到这些动物身上出现的问题,从而理解迁移体的生理功能。当然,在此之前,我们首先要知道哪些基因是形成迁移体所必需的。因此,我们将研究聚焦在研究迁移体的遗传通路和构建迁移体的体内模型上。在这两个目标之外,我们还需要开发研究迁移体的工具,特别是检测迁移体的方法和探针。
已 知
The Knowns
我们现在只需要使用GFP标记的迁移体标志物,例如PH-GFP,就可以轻松地在显微镜下检测到迁移体。此外,我们发现小麦精子蛋白WGA可以很好地将迁移体染色;基于这一发现,我们开发了一项使用WGA来检测迁移体的非常方便的方法。
迁移体形成时,伸缩丝从迁移细胞的轨迹边缘被拽出,这意味着伸缩丝必须粘附在细胞外基质上。由于整合素(integrin)是粘附于细胞外基质的主要分子,同时我们的质谱分析结果也显示迁移体上富集着整合素,因此我们将目光聚焦在整合素上。研究显示,具有活性的整合素在迁移体形成前就已经集合在了收缩丝的端中,活性整合素复合物和细胞外基质互相作用,沿着收缩丝搭建了多个粘附位点,这些位点随后成为迁移体形成的位点(Wu et al., 2017)。
目前,迁移体机制的研究进展大都起源于我们对迁移体形成过程的细胞生物学研究。鉴定出Tspan4是迁移体标志物后不久,我们还注意到,Tspan4似乎可以促进迁移体形成。Tspan4属于四跨膜蛋白家族,该家族有33个成员蛋白。我们发现,其中14个成员蛋白在过量表达的情况下都可以促进迁移体形成(Huang et al., 2019)。所有的四跨膜蛋白都有4个跨膜结构域,这些蛋白可以形成所谓的四跨膜蛋白富集的微结构域(TEM)(Rubinstein, 2011)。TEM尺寸大约为100纳米,富含一系列蛋白和脂阀类脂质,如胆固醇等。我们发现,在迁移体形成过程中,很多小的TEM会聚集形成微米级别的宏结构域,我们称之为四跨膜蛋白富集的宏结构域(TEMAs)。有意思的是,我们发现TEMA的形成与迁移体在收缩丝上的生长有关。这提示我们,TEMA的形成可能促使了收缩丝发生膜形变而形成迁移体。但是,我们怎么才能验证这个想法呢?
实验的关键进展源于一个意外的发现。在项目初期,我们计划使用体外系统来研究Tspan4在膜上的行为。因此,我们需要将Tspan4-GFP导入巨型单层脂质体(Giant Unilamellar Vesicle, GUV)中,观察Tspan4-GFP在膜上的行为。于是,我们首先把Tspan4整合到了蛋白脂质体中,随后通过电击融合蛋白脂质体来制备GUV。结果令人大吃一惊——在Tspan4-GFP蛋白脂质体电击融合的过程中,我们观察到了一些串珠状的结构,与迁移体及收缩丝的结构惊人的相似;而在对照组的蛋白脂质体的融合过程中却没有出现这样的结构。这个发现提示,Tspan4可能自己就足以驱动迁移体的形成。然而,由于我们并不清楚电击融合过程中究竟发生了什么,因此我们决定设计一个体外系统,来模拟迁移体的形成过程。
指导我们设计体外系统的关键灵感是我们意识到迁移细胞产生的拉力是迁移体产生的关键因素。因此,我们设计了两套不同的方案来将GUV拉成为膜管,以模拟收缩丝形成的过程。在第一套方案中,我们把GUV粘在流体小室中,然后使用液流推动GUV从而制造拉力。在第二套方案中,我们使用玻璃针头手动从GUV拉出膜管。这两套方案都让我们成功地从镶嵌了Tspan4-GFP的GUV中拉出膜管,在拉管过程中,TEM聚集形成TEMA,并在膜管上自发膨胀形成类似于迁移体的结构。
但是,为何TEMA的聚集会导致膜管膨胀成迁移体呢?科研中最让人难受的事莫过于做了一个完美的实验却无法解释其结果。万幸的是,我与以色列特拉维夫大学(Tel Aviv University)的迈克尔·科兹洛夫(Michael Kozlov)教授相识多年,他是个物理学出身的生物物理学家,是膜塑形领域的顶尖专家。迈克尔教授和他实验室一位聪明的研究生本·祖克(Ben Zuker)很快为收缩丝到迁移体的膜形变过程建立一个数学模型。该模型表明,当组合出的Tspan4富集的宏结构域的弯曲刚性超过膜的其余部位,收缩丝会自发地膨胀形成迁移体。这个模型可以用下面的类比来理解:假设我们有一个直径恒定的橡皮筋,但是不同部位的弹性不一样,当我们拉伸橡皮筋时,拉力会把橡皮筋拉长并拉细,硬的部位相比柔软的部位拉伸得少,因此会更粗。这个模型预测,TEMA的硬度会是收缩丝其余部位的5-10倍大, 在后续工作中, 原子力显微镜直接测量了迁移体膜的硬度,证实了这一预测。因此,迁移体的形成是由简单的物理过程所驱动的。
这一模型为我们提供了一个理解迁移体形成的理论框架。更重要的是,它确定了四跨膜蛋白在迁移体形成过程中的中心地位,此基础上,我们得以在研究迁移体的生理功能。
在研究迁移体和迁移性胞吐的生理功能时,最重要且最困难的部分是建立合适的动物模型。当时我们并不知道迁移体会在什么时间、什么部位、在哪种动物里出现。为了让搜索更加高效,我们制定了几条衡量好模型的标准。首先,模型必须便于显微镜观察。其次,模型需要容易开展遗传操作。最后,我们想要的模型应该是很容易获得的——如果隔壁实验室就有,那就再好不过了。最后,我们决定尝试斑马鱼胚胎。斑马鱼胚胎是透明的,因此是显微镜下研究的绝佳之选;而且基因敲除鱼很容易得到;最重要的是,使用斑马鱼为模型的著名发育生物学家孟安明教授就在我们隔壁楼里。
我们首先构建了表达迁移体标记蛋白的斑马鱼胚胎,并用活细胞成像技术观察这些胚胎。第一次试验中,我们就在活体胚胎里观测到了大量的迁移体和收缩丝(Jiang et al., 2019)。我们发现迁移体主要形成于原肠运动期。我们已经知道了迁移体形成所必需的一系列基因,如itgb1b, tspan4a 和tspan7,因此我们制备了敲除上述三个基因的斑马鱼。我们发现,迁移体的数量在敲除鱼的胚胎中有所减少,更有趣的是,这些迁移体减少的斑马鱼都出现了器官形成的缺陷——最明显的表型是不对称发育缺陷,包括左右翻转,多种器官的双侧复制(译注:即本该只在一侧出现的器官长在了身体另一侧或双侧都有)。
但这是否意味着敲除斑马鱼中的器官生成缺陷,是迁移体形成减少所致的呢?
答案是:并不一定。因为这些基因还具有与迁移体无关的其他功能,也可能是那些功能调控了器官生成。这样说来,迁移体形成也完全可能与器官生成毫不相关。实际上,在很多领域里,当我们去研究某个特定的细胞内事件的生理功能时,都会遇到类似的问题,至今也没有解决的办法。好在我们可以纯化迁移体,开展回补实验(rescue)——从野生型斑马鱼胚胎里纯化出迁移体,随后注射到敲除鱼的胚胎中。回补实验显示,敲除鱼的胚胎在注射了迁移体后,成功地恢复了器官形成,这表明器官形成的确依赖于迁移体,而不是Tspan4和Tspan7基因中那些与迁移体无关的其他功能。
下一个问题,迁移体又是如何调控器官生成的呢?我们推断迁移体可能包含某些信号分子,这些分子会在器官生成过程中指导细胞做出正确响应。为了检测这个理论,我们使用定量质谱的方法分析了纯化的迁移体。我们发现,迁移体中的确富集了一批配体分子,如趋化因子、细胞因子、形态发生素、生长因子等。其中,我们特别注意到了趋化因子CXCL12,因为CXCL12a的敲除鱼出现了和迁移体缺陷鱼相似的不对称发育缺陷。此外,将野生斑马鱼胚胎中的迁移体注射入CXCL12a敲除的斑马鱼胚胎中,能够成功恢复不对称发育的缺陷。因此,我们认为CXCL12是迁移体调控不对称发育中的关键分子。
在斑马鱼胚胎中,有一种带纤毛的器官叫库氏泡(Kupffer’s vesicle,KV),它能导致其他器官的左右不对称发育。我们发现,迁移体对KV的形成是必需的:在迁移体缺陷的胚胎中,KV不能有效形成。KV由一组迁移性前体细胞发育而来,这组细胞被称为背部先驱细胞(dorsal forerunner cells, DFCs)。在迁移体缺陷胚胎中,DFCs在迁移过程中跑散了,无法到达它们的目的地。这提示了一种可能性,即,迁移体作为区域性的趋化信号,指引迁移型DFC到达胚胎正确位置。为验证这一假说,我们从野生型胚胎中纯化得到迁移体,把它们嵌入琼脂糖珠上,随后将这些珠子插入胚胎的腹部一侧(正常情况下DFCs不会在腹部出现)。我们发现,含有迁移体的珠子可以吸引DFCs到达腹部。因此确认,迁移体是DFCs的趋化信号。
现在,拼图还剩最后一块没有完成。如果迁移体是DFCs的趋化信号,它们一定需要在DFCs迁移路径的终点区域富集。目前的研究已证实DFCs会在胚盾(译注:即embryonic shield,脊椎动物胚胎发育初期出现的不透明盾牌样结构)富集;因此我们检查了胚盾周围的区域。让我们意外的是,我们在胚盾下方发现了一个大空腔,并称之为“胚盾腔”(embryonic shield cavity)。迁移体在这个腔体内高度富集;当我们向斑马鱼胚胎注射纯化的迁移体时,它们最终会停留在胚盾腔里。这也可以解释我们的补救实验为何能够成功。
以上这些工作确立了迁移体首个已知的生理功能,更重要的是,它提供了一个范例,可用来研究迁移体如何在其他生命过程中起作用。在这个范例中,信号分子被打包装入迁移体中,随后由迁移体在指定地点释放,从而激活周围的细胞。从这个意义上来说,迁移体可以整合空间、时间和特定化学信息,这些信息对协调胚胎发育等复杂生命过程中的细胞群体行为不可或缺。简而言之,迁移体就是带寄送地址的信息包裹。
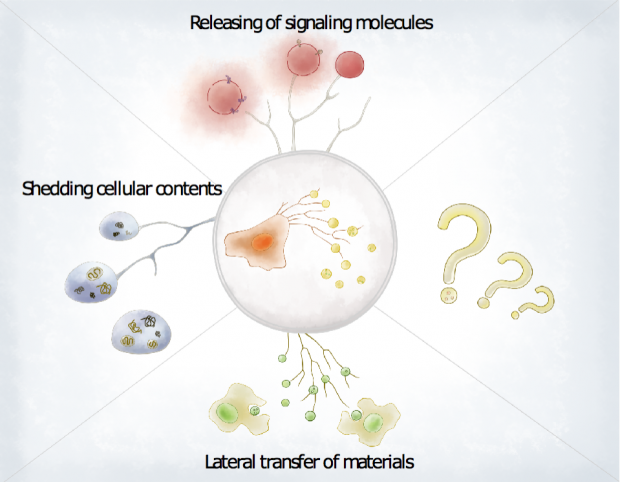
图2:迁移体的功能。
待解谜题
The Known Unknowns
从我们的研究延伸开去,还存着很多明显的问题有待回答。此处总结如下,并分享一些我的想法。
1)迁移体的生物发生机制究竟是什么?
当前,除了我们鉴定到的少数分子,控制迁移体形成的信号通路仍不为人知。甚至,我们对迁移体的众多基本特点仍不够理解。例如,迁移体内部的囊泡是什么?这些囊泡是如何进入迁移体的?它们有何功能?
2)迁移体水平传递是否有生理功能?
我们知道迁移体能够被周围细胞吞掉,这可能是细胞之间物质传递的一种机制。最近,我们发现迁移体中会选择性地富集一些种类的mRNA,当迁移体被周围细胞吞掉后,迁移体中的mRNA会在吞掉迁移体的细胞中翻译成蛋白,并改变该细胞的行为 (Zhu et al, Cell Research, accepted)。当前,我们还不清楚迁移体除mRNA和蛋白外还可以传递哪些物质,也不清楚这种细胞间物质传递是否在体内发生并具有生理功能。
3)除了胚胎发育,迁移体还在哪些生物过程中起到重要作用?
很多免疫细胞在发育和免疫应答过程中会发生迁移,并分泌大量信号分子。这听上去就像是迁移体可以大展身手的场景。
4)迁移体有细胞自主性的功能吗?
细胞可否使用迁移体来吐出不想保留的物质?这样可以维持自身稳态、丢弃受损或有害的细胞成分、维持表面分子数目的最优水平……
5)迁移体的进化起源是什么?
它们来源于进化早期的单细胞生物,还是在多细胞生物出现后才出现?使用动物界内早期分支的模式生物系统,或许能够帮助我们回答这个有意思的问题。
未知的未知
Unknown Unknowns
科学史提醒我们,在我们能想到的待解之谜外,总有“未知的”未知——凭我们现有的知识,根本无法预料将来还会遇到哪些问题。面对这些未知的未知,我们必须承认,大自然总是比我们想的更复杂。我们要有一颗敢于不断挑战已有模式的心,也需要有一双锐利的眼睛,能挑出现有理论框架下无法解释的现象。怀着这样的心态,或许再加上点好运,我们终将开启满是未知却新奇的世界。
参考文献
1. Chen, L., Ma, L., and Yu, L. (2019). WGA is a probe for migrasomes. Cell Discovery 5.
2. Huang, Y., Zucker, B., Zhang, S., Elias, S., Zhu, Y., Chen, H., Ding, T., Li, Y., Sun, Y., Lou, J., et al. (2019). Migrasome formation is mediated by assembly of micron-scale tetraspanin macrodomains. Nat Cell Biol 21, 991-1002.
3. Jiang, D., Jiang, Z., Lu, D., Wang, X., Liang, H., Zhang, J., Meng, Y., Li, Y., Wu, D., Huang, Y., et al. (2019). Migrasomes provide regional cues for organ morphogenesis during zebrafish gastrulation. Nat Cell Biol 21, 966-977.
4. Ma, L., Li, Y., Peng, J., Wu, D., Zhao, X., Cui, Y., Chen, L., Yan, X., Du, Y., and Yu, L. (2015). Discovery of the migrasome, an organelle mediating release of cytoplasmic contents during cell migration. Cell Res 25, 24-38.
5. Rubinstein, E. (2011). The complexity of tetraspanins. Biochem Soc Trans 39, 501-505.
6. Taylor, A.C., and Robbins, E. (1963). Observations on microextensions from the surface of isolated vertebrate cells. Dev Biol 6, 660-673.
7. Wu, D., Xu, Y., Ding, T., Zu, Y., Yang, C., and Yu, L. (2017). Pairing of integrins with ECM proteins determines migrasome formation. Cell Res 27, 1397-1400.
8. Zhao, X., Lei, Y., Zheng, J., Peng, J., Li, Y., Yu, L., and Chen, Y. (2019). Identification of markers for migrasome detection. Cell Discov 5, 27.
话题:

0
推荐
财新博客版权声明:财新博客所发布文章及图片之版权属博主本人及/或相关权利人所有,未经博主及/或相关权利人单独授权,任何网站、平面媒体不得予以转载。财新网对相关媒体的网站信息内容转载授权并不包括财新博客的文章及图片。博客文章均为作者个人观点,不代表财新网的立场和观点。
 京公网安备 11010502034662号
京公网安备 11010502034662号 {kind=link}